Download as pdf or txt
You might also like
- Agarwood - Science Behind The Fragrance-Springer Singapore (2016)Document175 pagesAgarwood - Science Behind The Fragrance-Springer Singapore (2016)OsamaAliMoussa100% (2)
- 11 Chapter 2Document203 pages11 Chapter 2Rajashekhar D. B.No ratings yet
- Biderman's Chart of Coercion: Method Effect and Purpose VariantsDocument1 pageBiderman's Chart of Coercion: Method Effect and Purpose VariantsmariavillaresNo ratings yet
- Virus-Like ParticlesDocument7 pagesVirus-Like ParticlesmariavillaresNo ratings yet
- 2009-Comparative Sensitivity of PCR Primer Sets For Detection of Cryptosporidium ParvumDocument6 pages2009-Comparative Sensitivity of PCR Primer Sets For Detection of Cryptosporidium ParvumWiwien HendrawanNo ratings yet
- Articulo 8 CovidDocument33 pagesArticulo 8 CovidJuan OrlandoniNo ratings yet
- β-actin promoter: a potential therapeutic toolDocument12 pagesβ-actin promoter: a potential therapeutic tooladamos1945No ratings yet
- Genomics of The Severe Isolate of Maize Chlorotic: Dwarf VirusDocument7 pagesGenomics of The Severe Isolate of Maize Chlorotic: Dwarf VirusBelisa SaitoNo ratings yet
- Nucleic: Polymerase Chain Reaction Amplification Acid From Stool SpecimensDocument7 pagesNucleic: Polymerase Chain Reaction Amplification Acid From Stool SpecimensRabiatul 'raney' AdawiyahNo ratings yet
- ActivityDocument10 pagesActivityMarisela Sanchez ChNo ratings yet
- Benh VirusDocument7 pagesBenh VirusThu Hien NguyenNo ratings yet
- J. Virol.-2012-Cadena-Nava-3318-26Document9 pagesJ. Virol.-2012-Cadena-Nava-3318-26Marcos LoredoNo ratings yet
- Ykkd GeneDocument8 pagesYkkd GeneKaviyarasu VelmuruganNo ratings yet
- Wang Et Al. - 2015 - Cloning and Iron Transportation of Nucleotide Binding Domain of Cryptosporidium Andersoni ATP-Binding Cassette (CaADocument6 pagesWang Et Al. - 2015 - Cloning and Iron Transportation of Nucleotide Binding Domain of Cryptosporidium Andersoni ATP-Binding Cassette (CaAWiwien HendrawanNo ratings yet
- Assignment Namin Ni MaderDocument6 pagesAssignment Namin Ni MaderGervy Racoma ValerosoNo ratings yet
- TMP 882 FDocument6 pagesTMP 882 FFrontiersNo ratings yet
- Argentina Beta ADocument3 pagesArgentina Beta Aelizabel andradeNo ratings yet
- Cloning, Characterization, and Distribution of An mRNA Encoding A H + - ATPase Subunit in The Mantle of Pearl Oyster, Pinctada FucataDocument8 pagesCloning, Characterization, and Distribution of An mRNA Encoding A H + - ATPase Subunit in The Mantle of Pearl Oyster, Pinctada FucataGunawan MuhammadNo ratings yet
- CH 09 PDFDocument17 pagesCH 09 PDFSyed Ali Akbar BokhariNo ratings yet
- Tyrosine: Receptor Phosphatase Mediates Homophilic BindingDocument9 pagesTyrosine: Receptor Phosphatase Mediates Homophilic BindingLUCAS GONÇALVES DE OLIVEIRANo ratings yet
- Janke Et Al 2004 AaYeastDocument16 pagesJanke Et Al 2004 AaYeastTolga Tarkan ÖlmezNo ratings yet
- Shipman2017 PDFDocument14 pagesShipman2017 PDFError404fucknicknameNo ratings yet
- Single Plasmid Systems For Inducible Dual ProteinDocument12 pagesSingle Plasmid Systems For Inducible Dual ProteinGregorius HocevarNo ratings yet
- Nucleotide Sequence Organization: and Genome of Canine ParvovirusDocument11 pagesNucleotide Sequence Organization: and Genome of Canine Parvovirushappycold00No ratings yet
- Dna 2009 0897Document10 pagesDna 2009 0897Ujwal TrivediNo ratings yet
- 16S Ribosomal DNA For: Amplification Phylogenetic StudyDocument7 pages16S Ribosomal DNA For: Amplification Phylogenetic StudyolamicroNo ratings yet
- 1 s2.0 S002075190800180X MainDocument10 pages1 s2.0 S002075190800180X Mainyujiao MaNo ratings yet
- Weisburg Et Al 1991 16s Ribosomal Dna Amplification For Phylogenetic StudyDocument7 pagesWeisburg Et Al 1991 16s Ribosomal Dna Amplification For Phylogenetic Studyteacher.mireyaNo ratings yet
- 28 Pgen.1001344Document11 pages28 Pgen.1001344Lucia UriasNo ratings yet
- LeviDocument2 pagesLeviAyioKunNo ratings yet
- Weisburg Et Al 1991 16s Ribosomal Dna Amplification For Phylogenetic StudyDocument7 pagesWeisburg Et Al 1991 16s Ribosomal Dna Amplification For Phylogenetic Studyteacher.mireyaNo ratings yet
- Ncomms 15464Document10 pagesNcomms 15464نوره نورNo ratings yet
- Pone 0020505Document9 pagesPone 0020505Monserrat EspinosaNo ratings yet
- Article1380286349 - Lan Et AlDocument6 pagesArticle1380286349 - Lan Et AlsuryaNo ratings yet
- 5-Microbiology Resource Announcements - 2019 - Val-Calvo J Et, AlDocument2 pages5-Microbiology Resource Announcements - 2019 - Val-Calvo J Et, AlAntonNo ratings yet
- J. Clin. Microbiol.-2011-Wolfaardt-728-31Document4 pagesJ. Clin. Microbiol.-2011-Wolfaardt-728-31NengLukmanNo ratings yet
- Yac Bac PacDocument30 pagesYac Bac PacRahmama100% (1)
- PIIS1674205217300047Document3 pagesPIIS1674205217300047Alberto Luis Lizcano GonzálezNo ratings yet
- Abe 2011Document9 pagesAbe 2011Shima YousefiNo ratings yet
- Lecture 1Document38 pagesLecture 1Đức Huy NguyễnNo ratings yet
- A Sea Urchin Genome Project: Sequence Scan, Virtual Map, and Additional ResourcesDocument5 pagesA Sea Urchin Genome Project: Sequence Scan, Virtual Map, and Additional ResourcesPradip ChauhanNo ratings yet
- Plat-E: An Efficient and Stable System For Transient Packaging of RetrovirusesDocument5 pagesPlat-E: An Efficient and Stable System For Transient Packaging of RetrovirusesSairam06No ratings yet
- Enhanced Foreign Protein Accumulation in Nicotiana Benthamiana Leaves Co-Infiltrated With A TMV Vector and Plant Cell Cycle Regulator GenesDocument7 pagesEnhanced Foreign Protein Accumulation in Nicotiana Benthamiana Leaves Co-Infiltrated With A TMV Vector and Plant Cell Cycle Regulator Geneszhaoyue12112001No ratings yet
- Detection and Analysis of Circular RNAs by RT-PCRDocument9 pagesDetection and Analysis of Circular RNAs by RT-PCRBahlibiNo ratings yet
- BGi RNA-Seq AnalysisDocument19 pagesBGi RNA-Seq Analysisshashikanth_marriNo ratings yet
- Content ServerDocument11 pagesContent ServerKeri Gobin SamarooNo ratings yet
- Plasmodium Vivax: Parasitemia Determination by Real-Time Quantitative PCR in Aotus MonkeysDocument4 pagesPlasmodium Vivax: Parasitemia Determination by Real-Time Quantitative PCR in Aotus MonkeysAndres JarrinNo ratings yet
- 2002-Specific Bovine Antibody Response Against A New Recombinant Cryptosporidium Parvum Antigen Containing 4 Zinc-Finger MotifsDocument6 pages2002-Specific Bovine Antibody Response Against A New Recombinant Cryptosporidium Parvum Antigen Containing 4 Zinc-Finger MotifswiwienNo ratings yet
- pBAD - IPTGDocument5 pagespBAD - IPTGMouna RohanaNo ratings yet
- Journal of Virology-1993-Langeveld-765.fullDocument8 pagesJournal of Virology-1993-Langeveld-765.fullSkuukzky baeNo ratings yet
- Preston1991Document5 pagesPreston1991Alberto TerronesNo ratings yet
- Artigo - 6 - Cardinali-Rezende - Genome Announc.-2015-Cardinali-Rezende PDFDocument2 pagesArtigo - 6 - Cardinali-Rezende - Genome Announc.-2015-Cardinali-Rezende PDFJuliana Cardinali RezendeNo ratings yet
- Disentangling Primer Interactions Improves SARS-CoV-2 Genome Sequencing by Multiplex Tiling PCRDocument11 pagesDisentangling Primer Interactions Improves SARS-CoV-2 Genome Sequencing by Multiplex Tiling PCRJulio BarriosNo ratings yet
- Journal of Virology-2016-Lu-1345.fullDocument14 pagesJournal of Virology-2016-Lu-1345.fullRayhana NaomiNo ratings yet
- Forsberg Et Al 1994 Use of Transcriptional Fusions To Monitor Gene Expression A Cautionary TaleDocument5 pagesForsberg Et Al 1994 Use of Transcriptional Fusions To Monitor Gene Expression A Cautionary TaleKunal KumarNo ratings yet
- CDU BIOINFORMATICS The Central DogmaDocument6 pagesCDU BIOINFORMATICS The Central DogmaKrisha Mae VillanuevaNo ratings yet
- Real Time PCRDocument9 pagesReal Time PCRGerson MouraNo ratings yet
- Cassette VDocument5 pagesCassette VnehamunjalNo ratings yet
- NorC, A New Efflux Pump Regulated by MgrA of Staphylococcus AureusDocument4 pagesNorC, A New Efflux Pump Regulated by MgrA of Staphylococcus Aureusamanda.thaisNo ratings yet
- Long Inverse PCRDocument4 pagesLong Inverse PCRdnajenNo ratings yet
- Aav Capsid Structure and Cell Interactions: Mavis Agbandje-Mckenna and Jürgen KleinschmidtDocument46 pagesAav Capsid Structure and Cell Interactions: Mavis Agbandje-Mckenna and Jürgen Kleinschmidtgauri ThapaNo ratings yet
- Genome Res.-1992-Ohler-51-9Document10 pagesGenome Res.-1992-Ohler-51-9Đặng Gia HoàngNo ratings yet
- DNA Methods in Food Safety: Molecular Typing of Foodborne and Waterborne Bacterial PathogensFrom EverandDNA Methods in Food Safety: Molecular Typing of Foodborne and Waterborne Bacterial PathogensNo ratings yet
- Leyendas BibDocument6 pagesLeyendas BibmariavillaresNo ratings yet
- Players AtlanticstormDocument16 pagesPlayers AtlanticstormmariavillaresNo ratings yet
- Fabian Society: Annual Report 2019/2020Document35 pagesFabian Society: Annual Report 2019/2020mariavillaresNo ratings yet
- Adam Weishaupt Human Devil PDFDocument53 pagesAdam Weishaupt Human Devil PDFxpat16391No ratings yet
- Salicylates and PandemiaDocument6 pagesSalicylates and PandemiamariavillaresNo ratings yet
- Operation Trust 1921 1926Document12 pagesOperation Trust 1921 1926mariavillaresNo ratings yet
- Bacterial Pneumonia Caused Most Deaths in 1918 Influenza PandemicDocument3 pagesBacterial Pneumonia Caused Most Deaths in 1918 Influenza Pandemicmariavillares100% (1)
- Virus-Like ParticlesDocument7 pagesVirus-Like ParticlesmariavillaresNo ratings yet
- WW1 and DisseasesDocument9 pagesWW1 and DisseasesmariavillaresNo ratings yet
- Against InfluenzaDocument13 pagesAgainst InfluenzamariavillaresNo ratings yet
- Research Progress: Bats in China Carry All The Ingredients To Make A New SARS VirusDocument2 pagesResearch Progress: Bats in China Carry All The Ingredients To Make A New SARS VirusmariavillaresNo ratings yet
- TrabajodelvirologochinoDocument4 pagesTrabajodelvirologochinomariavillaresNo ratings yet
- Vaccines CircaDocument10 pagesVaccines CircamariavillaresNo ratings yet
- Modification of Sars-Cov S1 Gene Render Expression in PichiaDocument18 pagesModification of Sars-Cov S1 Gene Render Expression in PichiamariavillaresNo ratings yet
- Agricultural Resource BaseDocument34 pagesAgricultural Resource BaseVenus AnteroNo ratings yet
- (Answer Key) Written Competency ReticulocytesDocument25 pages(Answer Key) Written Competency ReticulocytessaharNo ratings yet
- Organic ChemDocument116 pagesOrganic ChemNidhi Sisodia100% (1)
- LESLIE, C. Scientific Racism Reflections On Peer Review, Science and IdeologyDocument22 pagesLESLIE, C. Scientific Racism Reflections On Peer Review, Science and IdeologyÉden RodriguesNo ratings yet
- Population QuestionsDocument81 pagesPopulation QuestionsRiyan Moussaoui100% (1)
- List of Life SciencesDocument1 pageList of Life SciencesNathan Stuart The Retarded idiotNo ratings yet
- 16 - Chapter 12Document10 pages16 - Chapter 12Ritu ShewaniNo ratings yet
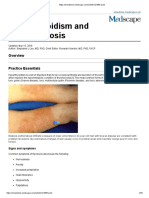
- Hyperthyroidism and ThyrotoxicosisDocument33 pagesHyperthyroidism and Thyrotoxicosiseze033No ratings yet
- Refractory Seizure in Childhood: Dyke-Davidoff-Masson Syndrome RevisitedDocument5 pagesRefractory Seizure in Childhood: Dyke-Davidoff-Masson Syndrome RevisitedaghniajolandaNo ratings yet
- Microbial Control: TH TH TH TH THDocument6 pagesMicrobial Control: TH TH TH TH THLaurence Bernard Jasper CanlasNo ratings yet
- Bile Metabolism and LithogenesisDocument15 pagesBile Metabolism and LithogenesisJEFFERSON ANDRES MUÑOZ LONDOÑONo ratings yet
- Epidemiology of Diabetes Mellitus PDFDocument425 pagesEpidemiology of Diabetes Mellitus PDFZulkifli Bakri Sallipadang100% (4)
- Chapter # 1: 1.1statement of The ProblemDocument12 pagesChapter # 1: 1.1statement of The ProblemShams Ud Din SahitoNo ratings yet
- 7-Pediatric - Drug TherapyDocument42 pages7-Pediatric - Drug TherapyDyn Adriani100% (1)
- Title: Gattaca: Guide QuestionsDocument3 pagesTitle: Gattaca: Guide Questionsbea andicoyNo ratings yet
- Anti-Aging Supplements & StrategiesDocument18 pagesAnti-Aging Supplements & StrategiesMateria by Purple AvenueNo ratings yet
- Antimicrobial Therapies: Methods and ProtocolsDocument436 pagesAntimicrobial Therapies: Methods and ProtocolsJanetteCardenasValdovinosNo ratings yet
- Biotransformation of DrugsDocument24 pagesBiotransformation of DrugsSteff SunshineNo ratings yet
- Pt. Pathlab Indonesia: (Laboratorium Klinik)Document1 pagePt. Pathlab Indonesia: (Laboratorium Klinik)Aghiest Utungga Al BirruNo ratings yet
- Nutrients 06 01782Document27 pagesNutrients 06 01782jasonc13No ratings yet
- AtaxiaDocument2 pagesAtaxiamohamed nagyNo ratings yet
- CancerDocument107 pagesCancervvenugopal74No ratings yet
- Drug-Induced Taste Disorders in Clinical Practice and Preclinical Safety EvaluationDocument10 pagesDrug-Induced Taste Disorders in Clinical Practice and Preclinical Safety EvaluationBharat NarumanchiNo ratings yet
- Leukemias & Lymphomas - HY USMLEDocument87 pagesLeukemias & Lymphomas - HY USMLEMatt McGlothlinNo ratings yet
- 통계 숙제 9Document2 pages통계 숙제 9Khongmuon NoiNo ratings yet
- Albrektsson Foreign Body ReactionDocument11 pagesAlbrektsson Foreign Body ReactionjeremyvoNo ratings yet
- Transgenic GuidelineDocument12 pagesTransgenic GuidelineYogesh BanginwarNo ratings yet
- Aspergillus SPDocument10 pagesAspergillus SPSandu PaulNo ratings yet