Download as pdf or txt
You might also like
- Coyotes Guide Chapter 3Document37 pagesCoyotes Guide Chapter 3Elan Goldbart100% (1)
- Child and Adolescents Prelim Exam Key To CorrectionDocument5 pagesChild and Adolescents Prelim Exam Key To CorrectionJosua RiveroNo ratings yet
- Life Sciences Fundamentals and Practice - IIDocument161 pagesLife Sciences Fundamentals and Practice - IIPathifnder Publication82% (22)
- Flora of Ethiopia and Eritrea - Aeonium 20200512 PDFDocument7 pagesFlora of Ethiopia and Eritrea - Aeonium 20200512 PDFCarles JiménezNo ratings yet
- Lungs & Gas Exchange Exam Questions: (Total 3 Marks)Document4 pagesLungs & Gas Exchange Exam Questions: (Total 3 Marks)Neelam HanifNo ratings yet
- Greek Latin Roots Revised For 4thDocument15 pagesGreek Latin Roots Revised For 4thNiki Eskew TurcoNo ratings yet
- EST II - BiologyDocument24 pagesEST II - BiologySaif100% (1)
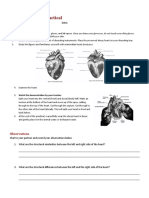
- Biology Worksheet Sheep Heart DissectionDocument3 pagesBiology Worksheet Sheep Heart DissectionFluphie Bunny0% (1)
- Plants: Unraveling The Phylogenomic Relationships of The Most Diverse African Palm Genus Raphia (Calamoideae, Arecaceae)Document20 pagesPlants: Unraveling The Phylogenomic Relationships of The Most Diverse African Palm Genus Raphia (Calamoideae, Arecaceae)Leandro RodríguezNo ratings yet
- FigueiredoEtMelo 2021 S NordestinensisDocument5 pagesFigueiredoEtMelo 2021 S NordestinensisgeorgesidneybarachoNo ratings yet
- Cinchoneae), A New Species From The Sira Mountains, Eastern Andes of Central Peru, and The Identity of Ladenbergia AcutifoliaDocument8 pagesCinchoneae), A New Species From The Sira Mountains, Eastern Andes of Central Peru, and The Identity of Ladenbergia AcutifoliaEdersin TorresNo ratings yet
- Chilquillo 2019Document6 pagesChilquillo 2019Edersin TorresNo ratings yet
- Pupulin Bogarin 2014 LepanthesguatemalensisgroupDocument18 pagesPupulin Bogarin 2014 LepanthesguatemalensisgroupHugo Rupérez PérezNo ratings yet
- Three New Lepanthes (Orchidaceae: Pleurothallidinae) From The Alto de Ventanas Ecoregion in Antioquia, ColombiaDocument14 pagesThree New Lepanthes (Orchidaceae: Pleurothallidinae) From The Alto de Ventanas Ecoregion in Antioquia, ColombiaSebastian VieiraNo ratings yet
- Rafflesia of The PhilippinesDocument17 pagesRafflesia of The Philippinesjo_dioNo ratings yet
- Taxonomy 04 00006 v2Document14 pagesTaxonomy 04 00006 v2ela.sofiaNo ratings yet
- A New Species of Renealmia (Zingiberaceae) From ColombiaDocument5 pagesA New Species of Renealmia (Zingiberaceae) From ColombiaRaúl PoznerNo ratings yet
- Flora MalaysiaDocument147 pagesFlora MalaysiaNadeesha GunasekaraNo ratings yet
- Geographic Range of Psychotria RubiaceaeDocument31 pagesGeographic Range of Psychotria RubiaceaeNicaRejusoNo ratings yet
- Neotropical MoraceaeDocument5 pagesNeotropical MoraceaeZin WadNo ratings yet
- Clave para Los Antoceros de Colombia PDFDocument9 pagesClave para Los Antoceros de Colombia PDFYennifer RamosNo ratings yet
- Description of Some Indian Veliidae in Different Zoogeographical Regions With Special Emphasis On Rhagovelia and MicroveliaDocument5 pagesDescription of Some Indian Veliidae in Different Zoogeographical Regions With Special Emphasis On Rhagovelia and MicroveliaKanhiya MahourNo ratings yet
- General Systematic Survey of Solanaceae and Convolvulaceae in Selected Philippine Plant RepresentativesDocument40 pagesGeneral Systematic Survey of Solanaceae and Convolvulaceae in Selected Philippine Plant RepresentativesJohn Rudolf Catalan100% (1)
- Espeletia RestrictaDocument7 pagesEspeletia RestrictaNatalia MontañoNo ratings yet
- Taxonomic Studies of Trees of The Cholistan Desert, PakistanDocument8 pagesTaxonomic Studies of Trees of The Cholistan Desert, PakistanInternational Network For Natural SciencesNo ratings yet
- Taxonomic Studies On The Family Solanaceae in The Rajshahi University CampusDocument6 pagesTaxonomic Studies On The Family Solanaceae in The Rajshahi University CampusOpenaccess Research paperNo ratings yet
- Chapter 2 Literature ReviewDocument30 pagesChapter 2 Literature ReviewSemana HumedalesNo ratings yet
- (Pollen Morphology of Neotropical Species of Podostemum (Malpighiales: Podostemaceae) )Document9 pages(Pollen Morphology of Neotropical Species of Podostemum (Malpighiales: Podostemaceae) )NilsonNo ratings yet
- The Genus Vanda (Orchidaceae: Aeridinae) in The Philippines: Two New Species and A Key To The Species in The ArchipelagoDocument10 pagesThe Genus Vanda (Orchidaceae: Aeridinae) in The Philippines: Two New Species and A Key To The Species in The ArchipelagoErik LegaspiNo ratings yet
- Rafflesia Aurantia Rafflesiaceae A New Species From PDFDocument12 pagesRafflesia Aurantia Rafflesiaceae A New Species From PDFJunegreg CualNo ratings yet
- Ghimire Et Al-2018-Nordic Journal of BotanyDocument30 pagesGhimire Et Al-2018-Nordic Journal of BotanyAndressa PelozoNo ratings yet
- PhcogRev 3 5 143Document11 pagesPhcogRev 3 5 143yermi atiNo ratings yet
- Flora of Peak Jabre, Paraíba, Brazilcactaceae - JussDocument6 pagesFlora of Peak Jabre, Paraíba, Brazilcactaceae - JussJohn GamesbyNo ratings yet
- 23 - 2022 - Devecchi Et Al. - Flora DuckeDocument12 pages23 - 2022 - Devecchi Et Al. - Flora DuckeMarcelo DevecchiNo ratings yet
- 2021c Clusia Falcata KewBullDocument6 pages2021c Clusia Falcata KewBullphyloNo ratings yet
- RubiaceaeDocument8 pagesRubiaceaex456456456xNo ratings yet
- 182644-Article Text-742690-7-10-20191119Document30 pages182644-Article Text-742690-7-10-20191119dindaNo ratings yet
- MCP 049Document11 pagesMCP 049rachiiidaNo ratings yet
- Anatomical Studies On The Genus Arabia (Subgenera:: Euphorbia L. Saudi Triucalli, Ermophyton, Esula and Chamaesyce)Document24 pagesAnatomical Studies On The Genus Arabia (Subgenera:: Euphorbia L. Saudi Triucalli, Ermophyton, Esula and Chamaesyce)etab alriashiNo ratings yet
- Checklist and Conservation Status of Ferns and Lycophytes in AmagDocument4 pagesChecklist and Conservation Status of Ferns and Lycophytes in AmagPerlita CaneteNo ratings yet
- Taxonomic Reevaluation of The Polylepis Sericea Complex (Rosaceae), With The Description of A New SpeciesDocument13 pagesTaxonomic Reevaluation of The Polylepis Sericea Complex (Rosaceae), With The Description of A New SpeciesMountains Forest MpedraceaeNo ratings yet
- Volum 1Document26 pagesVolum 1robertkellyyflNo ratings yet
- Denhametal CalyceraDocument25 pagesDenhametal Calycerahomies kunNo ratings yet
- Género CentrolobiumDocument16 pagesGénero CentrolobiumAnonymous ek4z9UBNo ratings yet
- Clave para Especies de Ludwigia L. (Onagraceae) de La Región Nor-Oriental eDocument5 pagesClave para Especies de Ludwigia L. (Onagraceae) de La Región Nor-Oriental eSMILSA MONTESINOS ROBLESNo ratings yet
- Iawa J 33 (2) 205-232Document28 pagesIawa J 33 (2) 205-232Phillip VinsonNo ratings yet
- Studies in Neotropical Vochysiaceae: A New Species of Vochysia (Section Ciliantha) From A Pluvial Forest of Western ColombiaDocument13 pagesStudies in Neotropical Vochysiaceae: A New Species of Vochysia (Section Ciliantha) From A Pluvial Forest of Western ColombiabioandreyNo ratings yet
- Lichens of New Zealand An Introductory Illustrated GuideDocument56 pagesLichens of New Zealand An Introductory Illustrated Guidecavris89% (9)
- Mendes-Silva Et Al. 2021 - Flora of Rondônia, Brazil Malmeeae (Annonaceae)Document27 pagesMendes-Silva Et Al. 2021 - Flora of Rondônia, Brazil Malmeeae (Annonaceae)Márcio BazanteNo ratings yet
- The Ant-Pollination System of Cytinus Hypocistis (Cytinaceae), A Mediterranean Root HoloparasiteDocument11 pagesThe Ant-Pollination System of Cytinus Hypocistis (Cytinaceae), A Mediterranean Root HoloparasiterachiiidaNo ratings yet
- Protolog PiperaceaeDocument40 pagesProtolog PiperaceaeFakhrana MeidaNo ratings yet
- HTTPSWWW - Nparks.gov - SG Mediasbgflora of Singaporevolume 13 Gentianales13, D, 1 Rubiaceae Lr1.AshxDocument358 pagesHTTPSWWW - Nparks.gov - SG Mediasbgflora of Singaporevolume 13 Gentianales13, D, 1 Rubiaceae Lr1.Ashxชนชนม์ ชูเชื้อNo ratings yet
- 27 1 117 TahilDocument8 pages27 1 117 TahilAbigail GarrinoNo ratings yet
- A New Species of Masdevallia (Orchidaceae: Pleurothallidinae) From PeruDocument1 pageA New Species of Masdevallia (Orchidaceae: Pleurothallidinae) From PeruEFRAIN SUCLLI MONTAÑEZNo ratings yet
- Vasco - Etal. - 2013 - Revision of Neotropical Elaphoglossum Squamipedia (DRYOP)Document44 pagesVasco - Etal. - 2013 - Revision of Neotropical Elaphoglossum Squamipedia (DRYOP)sandro.larosasanchezNo ratings yet
- Diversity and Distribution of Wild Edibl PDFDocument40 pagesDiversity and Distribution of Wild Edibl PDFZidni Ilman Navia M0% (1)
- TSWJ2014 150432Document15 pagesTSWJ2014 150432Anny LovibNo ratings yet
- MODULE 3 SeagrassDocument19 pagesMODULE 3 SeagrassMaria Rosalie NARITNo ratings yet
- Bryophytes (Liverworts) PDFDocument7 pagesBryophytes (Liverworts) PDFmanoj_rkl_07No ratings yet
- 25IJEAB 11020207 Identification PDFDocument6 pages25IJEAB 11020207 Identification PDFIJEAB JournalNo ratings yet
- A Paper On Cactaceae, Lobivia Ferox A New Record in Chilean FloraDocument8 pagesA Paper On Cactaceae, Lobivia Ferox A New Record in Chilean FloraJohn GamesbyNo ratings yet
- 3Document8 pages3Karina Winda BessufiNo ratings yet
- Almeda & Fidanza 2011 New SPP of Cambessedesia Harvard Papers Bot 16 57-63Document7 pagesAlmeda & Fidanza 2011 New SPP of Cambessedesia Harvard Papers Bot 16 57-63Giovanni Celso Paton MamaniNo ratings yet
- The Chilean Calceolaria Integrifolia Ehrhart2005Document30 pagesThe Chilean Calceolaria Integrifolia Ehrhart2005Evaldo PapeNo ratings yet
- DownloadDocument9 pagesDownloadEduarda SerraNo ratings yet
- Ollgaard - Windisch - 2016 - Lycopodiaceae in Brazil II. The Genera Lycopodiella, Palhinhaea and PseudolycopodiellaDocument29 pagesOllgaard - Windisch - 2016 - Lycopodiaceae in Brazil II. The Genera Lycopodiella, Palhinhaea and Pseudolycopodiellasandro.larosasanchezNo ratings yet
- Flora Zambesiaca Volume 13 Part 4: Xyridaceae, Eriocaulaceae, Typhaceae, Restionaceae, Flagellariaceae, Juncaceae, Musaceae, Strelitziaceae, Costaceae, Zingiberaceae, Cannaceae, MarantaceaeFrom EverandFlora Zambesiaca Volume 13 Part 4: Xyridaceae, Eriocaulaceae, Typhaceae, Restionaceae, Flagellariaceae, Juncaceae, Musaceae, Strelitziaceae, Costaceae, Zingiberaceae, Cannaceae, MarantaceaeNo ratings yet
- Taxonomic Position of Pinus Ceciliae Pinaceae EndeDocument10 pagesTaxonomic Position of Pinus Ceciliae Pinaceae EndeCarles JiménezNo ratings yet
- Typyfications of The Names of Iberian Accepted Species of Narcissus L. (Amaryllidaceae)Document10 pagesTypyfications of The Names of Iberian Accepted Species of Narcissus L. (Amaryllidaceae)Carles JiménezNo ratings yet
- Platanus ITS EvolutionDocument17 pagesPlatanus ITS EvolutionCarles JiménezNo ratings yet
- Cupressus Atlantica SEKIEWICZ - 2014 - 01Document19 pagesCupressus Atlantica SEKIEWICZ - 2014 - 01Carles JiménezNo ratings yet
- Amygdalus PhylogenyDocument17 pagesAmygdalus PhylogenyCarles JiménezNo ratings yet
- Genetic Diversity Wild Almonds PDFDocument21 pagesGenetic Diversity Wild Almonds PDFCarles JiménezNo ratings yet
- Can GuardiolaDocument18 pagesCan GuardiolaCarles JiménezNo ratings yet
- Pest and Pathogen Threats To Plane Trees 2015Document16 pagesPest and Pathogen Threats To Plane Trees 2015Carles JiménezNo ratings yet
- DNA Amounts MacaronesiaDocument12 pagesDNA Amounts MacaronesiaCarles JiménezNo ratings yet
- Platanus Orientalis CreticaDocument4 pagesPlatanus Orientalis CreticaCarles JiménezNo ratings yet
- Richardianus - Kirchnerianus 2019Document8 pagesRichardianus - Kirchnerianus 2019Carles JiménezNo ratings yet
- Jumping Through The Hoops BJLS 2019Document16 pagesJumping Through The Hoops BJLS 2019Carles JiménezNo ratings yet
- N Deficiens Herbert 1847Document7 pagesN Deficiens Herbert 1847Carles JiménezNo ratings yet
- Chasing Atlas DragonsDocument9 pagesChasing Atlas DragonsCarles JiménezNo ratings yet
- Article: A Natural Laboratory in Southern Spain: New Hybrids of Wild Daffodils (Narcissus, Amaryllidaceae)Document10 pagesArticle: A Natural Laboratory in Southern Spain: New Hybrids of Wild Daffodils (Narcissus, Amaryllidaceae)Carles JiménezNo ratings yet
- Narcissus - Vitekii Escobar - 2018Document10 pagesNarcissus - Vitekii Escobar - 2018Carles JiménezNo ratings yet
- Deficiens Túnez MEDAIL Et Al. 2020Document29 pagesDeficiens Túnez MEDAIL Et Al. 2020Carles JiménezNo ratings yet
- Genome Size Narcissus HybridsDocument9 pagesGenome Size Narcissus HybridsCarles JiménezNo ratings yet
- Amaryllideae Baker 1888Document5 pagesAmaryllideae Baker 1888Carles JiménezNo ratings yet
- Phylogeny AsparagalesDocument6 pagesPhylogeny AsparagalesCarles JiménezNo ratings yet
- Drimia Purpurascens Crespo Et Al 2020Document19 pagesDrimia Purpurascens Crespo Et Al 2020Carles JiménezNo ratings yet
- Aloe LukeanaDocument9 pagesAloe LukeanaCarles JiménezNo ratings yet
- Romano Et Al - Flora Vascolare Dell'isola Di Levanzo PDFDocument22 pagesRomano Et Al - Flora Vascolare Dell'isola Di Levanzo PDFCarles JiménezNo ratings yet
- Aloe Ikiorum ColeDocument6 pagesAloe Ikiorum ColeCarles JiménezNo ratings yet
- Drimia Anthericoides VELA - Et - Al - 2016 - PhytotaxaDocument8 pagesDrimia Anthericoides VELA - Et - Al - 2016 - PhytotaxaCarles JiménezNo ratings yet
- Mesoamerican Virola PDFDocument82 pagesMesoamerican Virola PDFCarles JiménezNo ratings yet
- GS COMPLETE For PRDocument246 pagesGS COMPLETE For PRHammad AHMEDNo ratings yet
- Seed Science and Technology1Document25 pagesSeed Science and Technology1DYCENo ratings yet
- Grasps in Science - BalangueDocument2 pagesGrasps in Science - BalangueBrielle Cailley BalangueNo ratings yet
- Genetics WebquestDocument2 pagesGenetics WebquestMariah ElieNo ratings yet
- Si in Fruit Crops FinalDocument49 pagesSi in Fruit Crops FinalRAKESH JANGIDNo ratings yet
- Punyashlok Ahilyadevi Holkar Solapur University, SolapurDocument17 pagesPunyashlok Ahilyadevi Holkar Solapur University, SolapurKalpak ShahaneNo ratings yet
- Objectives:: MLT 415 - Fundamentals of MicrobiologyDocument12 pagesObjectives:: MLT 415 - Fundamentals of MicrobiologyElujulo OyinkansolaNo ratings yet
- Intelligence - ReportDocument4 pagesIntelligence - ReportNyl FrayNo ratings yet
- MonsantoDocument33 pagesMonsantoakshit kiraneNo ratings yet
- Microbiology Fundamentals A Clinical Approach 2nd Edition Cowan Test BankDocument24 pagesMicrobiology Fundamentals A Clinical Approach 2nd Edition Cowan Test BankRodneyDanielsdnip97% (34)
- Evaa 155Document17 pagesEvaa 155Rogério MaruyamaNo ratings yet
- Sciencedirect Big Data Analytics For Personalized Medicine: Davide Cirillo and Alfonso ValenciaDocument10 pagesSciencedirect Big Data Analytics For Personalized Medicine: Davide Cirillo and Alfonso ValenciaAnyela Jim Alanya RamosNo ratings yet
- AERO200 Answer Key 9Document2 pagesAERO200 Answer Key 9Michael Yehezkiel RattuNo ratings yet
- APOCINACEAEDocument13 pagesAPOCINACEAEVenya Nabila VirdausyNo ratings yet
- Agr 203 Summary by Godspeed 3Document7 pagesAgr 203 Summary by Godspeed 3mongoldbreanna3No ratings yet
- PHD Thesis Structure BiologyDocument8 pagesPHD Thesis Structure BiologyWriteMyPaperCanadaSingapore100% (1)
- 2018 2019 Revised DVM Curriculum FinalDocument2 pages2018 2019 Revised DVM Curriculum FinalAguirre, John CastorNo ratings yet
- Nsejs Mock Test Student NoticeDocument1 pageNsejs Mock Test Student NoticeMuninder BhargavNo ratings yet
- Cte STD Principles Food ProductionDocument6 pagesCte STD Principles Food Productionryvnnnjovu13No ratings yet
- Mark Maslin - The Cradle of Humanity - How The Changing Landscape of Africa Made Us So Smart-Oxford University Press (2017)Document256 pagesMark Maslin - The Cradle of Humanity - How The Changing Landscape of Africa Made Us So Smart-Oxford University Press (2017)valeriu.ispir11No ratings yet
- Molecular Basis of InheritanceDocument5 pagesMolecular Basis of Inheritancesowmya gopinathNo ratings yet
- IB Biology Topic 7 - Nucleic Acids HL Revision SheetDocument1 pageIB Biology Topic 7 - Nucleic Acids HL Revision SheetFULYA YALDIZNo ratings yet
- Letm19 BiosciDocument16 pagesLetm19 Bioscigretelabelong10No ratings yet