
Mechanism of Action of Methotrexate in Rheumatoid Arthritis, and The Search For Biomarkers
Mechanism of Action of Methotrexate in Rheumatoid Arthritis, and The Search For Biomarkers
You might also like
- Goodness of God Key of EDocument2 pagesGoodness of God Key of Estephen2buizonNo ratings yet
- Research in Daily Life 2 Q1 Module 1Document22 pagesResearch in Daily Life 2 Q1 Module 1anggee100% (4)
- Classroom Supervisory Practices and Teacher Effectiveness Fil AuthorDocument13 pagesClassroom Supervisory Practices and Teacher Effectiveness Fil AuthorLAGUIO ELEMENTARY SCHOOLNo ratings yet
- Enzymology 1Document18 pagesEnzymology 1THE A plus ACADMYNo ratings yet
- Holdgate 2017Document18 pagesHoldgate 2017NofrizalNo ratings yet
- Reviews: Methotrexate and Its Mechanisms of Action in Inflammatory ArthritisDocument10 pagesReviews: Methotrexate and Its Mechanisms of Action in Inflammatory ArthritisanitaNo ratings yet
- Friedman 2018Document7 pagesFriedman 2018Genshin SatuNo ratings yet
- 1 s2.0 S1297319X18301817 MainDocument7 pages1 s2.0 S1297319X18301817 MainDanianto WicaksonoNo ratings yet
- Wang 2018Document42 pagesWang 2018Erni SalehNo ratings yet
- Will Pharmacogenetics Allow Better Prediction of MTX Toxicity and Efficacy in Patients With Rheumatoid ArthritisDocument7 pagesWill Pharmacogenetics Allow Better Prediction of MTX Toxicity and Efficacy in Patients With Rheumatoid Arthritisdoc0814No ratings yet
- MTX in D 2007Document13 pagesMTX in D 2007Ari KurniawanNo ratings yet
- Targeting Glutamate Signalling in Depression: Progress and ProspectsDocument15 pagesTargeting Glutamate Signalling in Depression: Progress and ProspectsCony GSNo ratings yet
- Polymorphism in Gene For ABCC2 Transporter PredictDocument12 pagesPolymorphism in Gene For ABCC2 Transporter PredictGregorius HocevarNo ratings yet
- Evolution of Macromolecular Complexity in Drug Delivery SystemsDocument17 pagesEvolution of Macromolecular Complexity in Drug Delivery SystemsAlessandroNo ratings yet
- Nrclinonc 2017 95Document13 pagesNrclinonc 2017 95김현수No ratings yet
- NRD 2016 211Document14 pagesNRD 2016 211김승훈(이과대학 화학)No ratings yet
- 2020 - Emerging Challenges To The Safe and Effective Use of Methadone For Cancer-Related Pain in Paediatric and Adult Patient PopulationsDocument16 pages2020 - Emerging Challenges To The Safe and Effective Use of Methadone For Cancer-Related Pain in Paediatric and Adult Patient PopulationsPoliana carmonaNo ratings yet
- Perspectives: Methotrexate-How Does It Really Work?Document4 pagesPerspectives: Methotrexate-How Does It Really Work?Genshin SatuNo ratings yet
- Understanding The Mechanisms of Action of MethotrexateDocument6 pagesUnderstanding The Mechanisms of Action of MethotrexateMuhamad Rizqy MaulanaNo ratings yet
- Association Between MTHFR C677T Polymorphism 2017 PDFDocument11 pagesAssociation Between MTHFR C677T Polymorphism 2017 PDFFaMe TanapornNo ratings yet
- Reviews: Proton-Pump Inhibitors: Understanding The Complications and RisksDocument14 pagesReviews: Proton-Pump Inhibitors: Understanding The Complications and RisksRONALD. D VIERA .MNo ratings yet
- Expanding The Medicinal Chemistry Synthetic ToolboxDocument19 pagesExpanding The Medicinal Chemistry Synthetic ToolboxPhoebeliza Jane BroñolaNo ratings yet
- Méthadone: de La Pharmacocinétique À La Pharmacologie CliniqueDocument9 pagesMéthadone: de La Pharmacocinétique À La Pharmacologie CliniqueBrasseurNo ratings yet
- Clinical Pharmacokinetics of Methotrexate: D.O. Shen and DL AzarnoffDocument2 pagesClinical Pharmacokinetics of Methotrexate: D.O. Shen and DL AzarnoffAdkhiatul MuslihatinNo ratings yet
- Biomedicines: Approaching Sites of Action of Temozolomide For Pharmacological and Clinical Studies in GlioblastomaDocument27 pagesBiomedicines: Approaching Sites of Action of Temozolomide For Pharmacological and Clinical Studies in GlioblastomaAtharva ThiteNo ratings yet
- A Patent Review of Arginine Methyltransferase Inhibitors (2010-2018)Document38 pagesA Patent Review of Arginine Methyltransferase Inhibitors (2010-2018)shamimNo ratings yet
- Supplementation With Folic Acid During Methotrexate Therapy For Rheumatoid ArthritisDocument9 pagesSupplementation With Folic Acid During Methotrexate Therapy For Rheumatoid ArthritisGenshin SatuNo ratings yet
- Regular ArticleDocument8 pagesRegular ArticleFaMe TanapornNo ratings yet
- Tahrani 2016Document27 pagesTahrani 2016Henry Leroy Lewis BatresNo ratings yet
- Molecular Therapeutics: ReviewDocument10 pagesMolecular Therapeutics: ReviewChika SabaNo ratings yet
- Membranosa 2Document17 pagesMembranosa 2Carolina CastroNo ratings yet
- Reviews: Marked For Death: Targeting Epigenetic Changes in CancerDocument23 pagesReviews: Marked For Death: Targeting Epigenetic Changes in CancerLorena RamosNo ratings yet
- Outcomes of Methotrexate Therapy For Psoriasis and Relationship To Genetic PolymorphismsDocument4 pagesOutcomes of Methotrexate Therapy For Psoriasis and Relationship To Genetic Polymorphismsizabela1980No ratings yet
- Methotrexate in Rheumatoid Arthritis Efficacy and Safety 2329 6887 2 127Document4 pagesMethotrexate in Rheumatoid Arthritis Efficacy and Safety 2329 6887 2 127Saifuddin HaswareNo ratings yet
- Methotrexate Side Effects Review Article 2157 7609.1000125Document5 pagesMethotrexate Side Effects Review Article 2157 7609.1000125Rania T. SabriNo ratings yet
- MacrophagesDocument18 pagesMacrophagesSaif WahidNo ratings yet
- Ac40c301 0f81 4a6e A47a 0440cb2a5ada 6020d3d73572f0002e215030 1612764156 The Changing Face of Pharmaceutical AnalysisDocument6 pagesAc40c301 0f81 4a6e A47a 0440cb2a5ada 6020d3d73572f0002e215030 1612764156 The Changing Face of Pharmaceutical AnalysisClarkStewartFaylogaErmilaNo ratings yet
- C1C13 and C14-Methotrexate - Methotrexate Sodium (CPhA Monograph)Document13 pagesC1C13 and C14-Methotrexate - Methotrexate Sodium (CPhA Monograph)Mina AzmyNo ratings yet
- Princ Farmacot Harrison PDFDocument13 pagesPrinc Farmacot Harrison PDFMade DeddyNo ratings yet
- Engineering and Physical Sciences in Oncology - Challenges and OpportunitiesDocument17 pagesEngineering and Physical Sciences in Oncology - Challenges and OpportunitiesAlessandroNo ratings yet
- Renal CellDocument10 pagesRenal CellEss liNo ratings yet
- Clinical Implications of PTEN Loss in Prostate CancerDocument13 pagesClinical Implications of PTEN Loss in Prostate Cancermedp7060No ratings yet
- Possibilities and Principle of MTXDocument9 pagesPossibilities and Principle of MTXPrillyPTNo ratings yet
- Micro TubuleDocument6 pagesMicro TubuleRntiaNo ratings yet
- 49.microbiota-Derived 3-IAA InfluencesDocument30 pages49.microbiota-Derived 3-IAA Influenceszhe zhNo ratings yet
- RA1Document4 pagesRA1Intan Winta PratiwiNo ratings yet
- Jashore University of Science and TechnologyDocument16 pagesJashore University of Science and TechnologyBlood DonationNo ratings yet
- Nature - Antithrombotic Therapy For PatientsDocument19 pagesNature - Antithrombotic Therapy For PatientsMatheus Augusto Salerno Miguel e SousaNo ratings yet
- Abdollahi-Roodsaz2016 Artigo3Document10 pagesAbdollahi-Roodsaz2016 Artigo3JamilaNo ratings yet
- Oligometastatic Prostate Cancer. Definitions, Clinical Outcomes, and Treatment Considerations (2017) .Document11 pagesOligometastatic Prostate Cancer. Definitions, Clinical Outcomes, and Treatment Considerations (2017) .Orlando SotoNo ratings yet
- The Journal of Rheumatology Volume 44, No. 6Document8 pagesThe Journal of Rheumatology Volume 44, No. 6FaMe TanapornNo ratings yet
- Primer: HypoparathyroidismDocument20 pagesPrimer: HypoparathyroidismKarenNo ratings yet
- Molecular Oncology - 2009 - Pierotti - Targeted Therapies The Rare Cancer ParadigmDocument19 pagesMolecular Oncology - 2009 - Pierotti - Targeted Therapies The Rare Cancer ParadigmJairo BermúdezNo ratings yet
- Methotrexate Alopecia AreataDocument10 pagesMethotrexate Alopecia AreataThorn MailNo ratings yet
- Methylene Blue in Anticancer Photodynamic TherapyDocument7 pagesMethylene Blue in Anticancer Photodynamic Therapyenriquecamacho.buyNo ratings yet
- Paper TranscripcionDocument13 pagesPaper TranscripcionKarime MontenegroNo ratings yet
- Kanji 2018Document6 pagesKanji 2018KEVIN ROGGER OBANDO BALLARDONo ratings yet
- Loss-Of-Function Genetic Tools For Animal Models - Cross-Species and Cross-Platform DifferencesDocument17 pagesLoss-Of-Function Genetic Tools For Animal Models - Cross-Species and Cross-Platform DifferencesLeon PalomeraNo ratings yet
- A Review of Methods To Determine Viability, Vitality, and Metabolic Rates in MicrobiologyDocument25 pagesA Review of Methods To Determine Viability, Vitality, and Metabolic Rates in MicrobiologyAna Carolina FurianNo ratings yet
- Paper HydrogelDocument17 pagesPaper HydrogelAntar GayenNo ratings yet
- 2016 Medical Ozone Increases Methotrexate Clinical Response and Improves Cellular Redox Balance in Patients With Rheumatoid ArthritisDocument6 pages2016 Medical Ozone Increases Methotrexate Clinical Response and Improves Cellular Redox Balance in Patients With Rheumatoid ArthritisAcupuntura de EquinosNo ratings yet
- JGM 2990Document7 pagesJGM 2990FaMe TanapornNo ratings yet
- Metabolite Safety in Drug DevelopmentFrom EverandMetabolite Safety in Drug DevelopmentSuzanne L. IversonNo ratings yet
- JGM 2990Document7 pagesJGM 2990FaMe TanapornNo ratings yet
- Pharmacokinetics and Pharmacogenetics of High Dose Methotrexate in Chinese Adult Patients With Non Hodgkin Lymphoma: A Population AnalysisDocument17 pagesPharmacokinetics and Pharmacogenetics of High Dose Methotrexate in Chinese Adult Patients With Non Hodgkin Lymphoma: A Population AnalysisFaMe TanapornNo ratings yet
- Key 275Document4 pagesKey 275FaMe TanapornNo ratings yet
- MTHFR Gene Polymorphisms and Susceptibility To Rheumatoid Arthritis: A Meta-Analysis Based On 16 StudiesDocument13 pagesMTHFR Gene Polymorphisms and Susceptibility To Rheumatoid Arthritis: A Meta-Analysis Based On 16 StudiesFaMe TanapornNo ratings yet
- Association Between MTHFR C677T Polymorphism 2017 PDFDocument11 pagesAssociation Between MTHFR C677T Polymorphism 2017 PDFFaMe TanapornNo ratings yet
- 1070 Impact of MTHFR rs1801133 MTHFR rs1801131 and Abcb1 rs1045642 Polymorphisms With Increased Susceptibility of Rheumatoid Arthritis in The West Algerian Population A Case Control Study FileDocument9 pages1070 Impact of MTHFR rs1801133 MTHFR rs1801131 and Abcb1 rs1045642 Polymorphisms With Increased Susceptibility of Rheumatoid Arthritis in The West Algerian Population A Case Control Study FileFaMe TanapornNo ratings yet
- Acr 23164Document11 pagesAcr 23164FaMe TanapornNo ratings yet
- Z37 PDFDocument8 pagesZ37 PDFFaMe TanapornNo ratings yet
- Regular ArticleDocument8 pagesRegular ArticleFaMe TanapornNo ratings yet
- Use of MTHFR C677T Polymorphism and Plasma Pharmacokinetics To Predict Methotrexate Toxicity in Patients With Acute Lymphoblastic LeukemiaDocument8 pagesUse of MTHFR C677T Polymorphism and Plasma Pharmacokinetics To Predict Methotrexate Toxicity in Patients With Acute Lymphoblastic LeukemiaFaMe TanapornNo ratings yet
- The Journal of Rheumatology Volume 44, No. 6Document8 pagesThe Journal of Rheumatology Volume 44, No. 6FaMe TanapornNo ratings yet
- Sciencedirect: Original ArticleDocument8 pagesSciencedirect: Original ArticleFaMe TanapornNo ratings yet
- Document Analysis Worksheet: Analyze A PaintingDocument4 pagesDocument Analysis Worksheet: Analyze A PaintingPatrick SanchezNo ratings yet
- Swapnil CromaDocument15 pagesSwapnil Cromapanpatil swapnilNo ratings yet
- Medicinal Benefits of HoneyDocument8 pagesMedicinal Benefits of HoneyTauseef AshrafNo ratings yet
- Zabala Auto SupplyDocument7 pagesZabala Auto SupplyLeighsen VillacortaNo ratings yet
- What Is The Difference Between Graduate and UndergraduateDocument6 pagesWhat Is The Difference Between Graduate and UndergraduateWinnie OchiengNo ratings yet
- CME Applique TutorialDocument20 pagesCME Applique TutorialMarittaKarmaNo ratings yet
- (1899) (Vol. 2) A Sailor's Life Under Four SovereignsDocument364 pages(1899) (Vol. 2) A Sailor's Life Under Four SovereignsHerbert Hillary Booker 2ndNo ratings yet
- ToninaDocument6 pagesToninapancholopez2014No ratings yet
- HZ2322 EM250 Series 100mW ZigBee Module (ENG)Document10 pagesHZ2322 EM250 Series 100mW ZigBee Module (ENG)MicroComSolutionsNo ratings yet
- Artigo Hambate BaDocument19 pagesArtigo Hambate BaRayza GiardiniNo ratings yet
- DAS AppFormDocument3 pagesDAS AppFormZia HashmiNo ratings yet
- Integrated NLP Train The Trainer BootcampDocument8 pagesIntegrated NLP Train The Trainer BootcampHitesh ParmarNo ratings yet
- Santiago v. Gonzalez PDFDocument2 pagesSantiago v. Gonzalez PDFGervin ArquizalNo ratings yet
- MSDS Abate - 1 - GR-ID-SDS - 02 - GHSDocument34 pagesMSDS Abate - 1 - GR-ID-SDS - 02 - GHSpurwadiNo ratings yet
- 22TCN263 2000 Tieu Chuan Khao Sat Duong o ToDocument109 pages22TCN263 2000 Tieu Chuan Khao Sat Duong o ToTu PhamNo ratings yet
- Vampires Werewolves and Witches Oh MyDocument51 pagesVampires Werewolves and Witches Oh Myangelg29850% (1)
- 2.2 - Concrete Cracked Elastic Stresses StageDocument12 pages2.2 - Concrete Cracked Elastic Stresses StageMarcelo AbreraNo ratings yet
- Guidelines On Good Research PracticeDocument9 pagesGuidelines On Good Research PracticeAndre SetiawanNo ratings yet
- Weaning Meal Plan 7 9 MonthsDocument1 pageWeaning Meal Plan 7 9 MonthsDeborah EvangelineNo ratings yet
- Dark Souls 3 Lore - Father Ariandel & Sister Friede - VaatividyaDocument5 pagesDark Souls 3 Lore - Father Ariandel & Sister Friede - VaatividyashaunNo ratings yet
- PKM Full C.V. Sept 2021Document11 pagesPKM Full C.V. Sept 2021aopera87No ratings yet
- Startup Law OverviewDocument11 pagesStartup Law OverviewAgibise AkanliseNo ratings yet
- VollyballDocument4 pagesVollyballCristina LataganNo ratings yet
- Final ExamDocument3 pagesFinal ExamFaidah PangandamanNo ratings yet
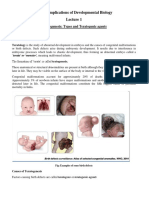
- Unit 5: Implications of Developmental Biology: Teratogenesis: Types and Teratogenic AgentsDocument12 pagesUnit 5: Implications of Developmental Biology: Teratogenesis: Types and Teratogenic AgentsAmar Kant JhaNo ratings yet
- Just Say No To Big Brother's Smart Meters: The Latest in Bio-Hazard Technology by Orlean KoehleDocument148 pagesJust Say No To Big Brother's Smart Meters: The Latest in Bio-Hazard Technology by Orlean KoehleVen GeanciaNo ratings yet
- Diagnosis of Stroke-Associated PneumoniaDocument16 pagesDiagnosis of Stroke-Associated PneumoniaCecilia Casandra UneputtyNo ratings yet































































































Download as pdf or txt
You might also like
- Goodness of God Key of EDocument2 pagesGoodness of God Key of Estephen2buizonNo ratings yet
- Research in Daily Life 2 Q1 Module 1Document22 pagesResearch in Daily Life 2 Q1 Module 1anggee100% (4)
- Classroom Supervisory Practices and Teacher Effectiveness Fil AuthorDocument13 pagesClassroom Supervisory Practices and Teacher Effectiveness Fil AuthorLAGUIO ELEMENTARY SCHOOLNo ratings yet
- Enzymology 1Document18 pagesEnzymology 1THE A plus ACADMYNo ratings yet
- Holdgate 2017Document18 pagesHoldgate 2017NofrizalNo ratings yet
- Reviews: Methotrexate and Its Mechanisms of Action in Inflammatory ArthritisDocument10 pagesReviews: Methotrexate and Its Mechanisms of Action in Inflammatory ArthritisanitaNo ratings yet
- Friedman 2018Document7 pagesFriedman 2018Genshin SatuNo ratings yet
- 1 s2.0 S1297319X18301817 MainDocument7 pages1 s2.0 S1297319X18301817 MainDanianto WicaksonoNo ratings yet
- Wang 2018Document42 pagesWang 2018Erni SalehNo ratings yet
- Will Pharmacogenetics Allow Better Prediction of MTX Toxicity and Efficacy in Patients With Rheumatoid ArthritisDocument7 pagesWill Pharmacogenetics Allow Better Prediction of MTX Toxicity and Efficacy in Patients With Rheumatoid Arthritisdoc0814No ratings yet
- MTX in D 2007Document13 pagesMTX in D 2007Ari KurniawanNo ratings yet
- Targeting Glutamate Signalling in Depression: Progress and ProspectsDocument15 pagesTargeting Glutamate Signalling in Depression: Progress and ProspectsCony GSNo ratings yet
- Polymorphism in Gene For ABCC2 Transporter PredictDocument12 pagesPolymorphism in Gene For ABCC2 Transporter PredictGregorius HocevarNo ratings yet
- Evolution of Macromolecular Complexity in Drug Delivery SystemsDocument17 pagesEvolution of Macromolecular Complexity in Drug Delivery SystemsAlessandroNo ratings yet
- Nrclinonc 2017 95Document13 pagesNrclinonc 2017 95김현수No ratings yet
- NRD 2016 211Document14 pagesNRD 2016 211김승훈(이과대학 화학)No ratings yet
- 2020 - Emerging Challenges To The Safe and Effective Use of Methadone For Cancer-Related Pain in Paediatric and Adult Patient PopulationsDocument16 pages2020 - Emerging Challenges To The Safe and Effective Use of Methadone For Cancer-Related Pain in Paediatric and Adult Patient PopulationsPoliana carmonaNo ratings yet
- Perspectives: Methotrexate-How Does It Really Work?Document4 pagesPerspectives: Methotrexate-How Does It Really Work?Genshin SatuNo ratings yet
- Understanding The Mechanisms of Action of MethotrexateDocument6 pagesUnderstanding The Mechanisms of Action of MethotrexateMuhamad Rizqy MaulanaNo ratings yet
- Association Between MTHFR C677T Polymorphism 2017 PDFDocument11 pagesAssociation Between MTHFR C677T Polymorphism 2017 PDFFaMe TanapornNo ratings yet
- Reviews: Proton-Pump Inhibitors: Understanding The Complications and RisksDocument14 pagesReviews: Proton-Pump Inhibitors: Understanding The Complications and RisksRONALD. D VIERA .MNo ratings yet
- Expanding The Medicinal Chemistry Synthetic ToolboxDocument19 pagesExpanding The Medicinal Chemistry Synthetic ToolboxPhoebeliza Jane BroñolaNo ratings yet
- Méthadone: de La Pharmacocinétique À La Pharmacologie CliniqueDocument9 pagesMéthadone: de La Pharmacocinétique À La Pharmacologie CliniqueBrasseurNo ratings yet
- Clinical Pharmacokinetics of Methotrexate: D.O. Shen and DL AzarnoffDocument2 pagesClinical Pharmacokinetics of Methotrexate: D.O. Shen and DL AzarnoffAdkhiatul MuslihatinNo ratings yet
- Biomedicines: Approaching Sites of Action of Temozolomide For Pharmacological and Clinical Studies in GlioblastomaDocument27 pagesBiomedicines: Approaching Sites of Action of Temozolomide For Pharmacological and Clinical Studies in GlioblastomaAtharva ThiteNo ratings yet
- A Patent Review of Arginine Methyltransferase Inhibitors (2010-2018)Document38 pagesA Patent Review of Arginine Methyltransferase Inhibitors (2010-2018)shamimNo ratings yet
- Supplementation With Folic Acid During Methotrexate Therapy For Rheumatoid ArthritisDocument9 pagesSupplementation With Folic Acid During Methotrexate Therapy For Rheumatoid ArthritisGenshin SatuNo ratings yet
- Regular ArticleDocument8 pagesRegular ArticleFaMe TanapornNo ratings yet
- Tahrani 2016Document27 pagesTahrani 2016Henry Leroy Lewis BatresNo ratings yet
- Molecular Therapeutics: ReviewDocument10 pagesMolecular Therapeutics: ReviewChika SabaNo ratings yet
- Membranosa 2Document17 pagesMembranosa 2Carolina CastroNo ratings yet
- Reviews: Marked For Death: Targeting Epigenetic Changes in CancerDocument23 pagesReviews: Marked For Death: Targeting Epigenetic Changes in CancerLorena RamosNo ratings yet
- Outcomes of Methotrexate Therapy For Psoriasis and Relationship To Genetic PolymorphismsDocument4 pagesOutcomes of Methotrexate Therapy For Psoriasis and Relationship To Genetic Polymorphismsizabela1980No ratings yet
- Methotrexate in Rheumatoid Arthritis Efficacy and Safety 2329 6887 2 127Document4 pagesMethotrexate in Rheumatoid Arthritis Efficacy and Safety 2329 6887 2 127Saifuddin HaswareNo ratings yet
- Methotrexate Side Effects Review Article 2157 7609.1000125Document5 pagesMethotrexate Side Effects Review Article 2157 7609.1000125Rania T. SabriNo ratings yet
- MacrophagesDocument18 pagesMacrophagesSaif WahidNo ratings yet
- Ac40c301 0f81 4a6e A47a 0440cb2a5ada 6020d3d73572f0002e215030 1612764156 The Changing Face of Pharmaceutical AnalysisDocument6 pagesAc40c301 0f81 4a6e A47a 0440cb2a5ada 6020d3d73572f0002e215030 1612764156 The Changing Face of Pharmaceutical AnalysisClarkStewartFaylogaErmilaNo ratings yet
- C1C13 and C14-Methotrexate - Methotrexate Sodium (CPhA Monograph)Document13 pagesC1C13 and C14-Methotrexate - Methotrexate Sodium (CPhA Monograph)Mina AzmyNo ratings yet
- Princ Farmacot Harrison PDFDocument13 pagesPrinc Farmacot Harrison PDFMade DeddyNo ratings yet
- Engineering and Physical Sciences in Oncology - Challenges and OpportunitiesDocument17 pagesEngineering and Physical Sciences in Oncology - Challenges and OpportunitiesAlessandroNo ratings yet
- Renal CellDocument10 pagesRenal CellEss liNo ratings yet
- Clinical Implications of PTEN Loss in Prostate CancerDocument13 pagesClinical Implications of PTEN Loss in Prostate Cancermedp7060No ratings yet
- Possibilities and Principle of MTXDocument9 pagesPossibilities and Principle of MTXPrillyPTNo ratings yet
- Micro TubuleDocument6 pagesMicro TubuleRntiaNo ratings yet
- 49.microbiota-Derived 3-IAA InfluencesDocument30 pages49.microbiota-Derived 3-IAA Influenceszhe zhNo ratings yet
- RA1Document4 pagesRA1Intan Winta PratiwiNo ratings yet
- Jashore University of Science and TechnologyDocument16 pagesJashore University of Science and TechnologyBlood DonationNo ratings yet
- Nature - Antithrombotic Therapy For PatientsDocument19 pagesNature - Antithrombotic Therapy For PatientsMatheus Augusto Salerno Miguel e SousaNo ratings yet
- Abdollahi-Roodsaz2016 Artigo3Document10 pagesAbdollahi-Roodsaz2016 Artigo3JamilaNo ratings yet
- Oligometastatic Prostate Cancer. Definitions, Clinical Outcomes, and Treatment Considerations (2017) .Document11 pagesOligometastatic Prostate Cancer. Definitions, Clinical Outcomes, and Treatment Considerations (2017) .Orlando SotoNo ratings yet
- The Journal of Rheumatology Volume 44, No. 6Document8 pagesThe Journal of Rheumatology Volume 44, No. 6FaMe TanapornNo ratings yet
- Primer: HypoparathyroidismDocument20 pagesPrimer: HypoparathyroidismKarenNo ratings yet
- Molecular Oncology - 2009 - Pierotti - Targeted Therapies The Rare Cancer ParadigmDocument19 pagesMolecular Oncology - 2009 - Pierotti - Targeted Therapies The Rare Cancer ParadigmJairo BermúdezNo ratings yet
- Methotrexate Alopecia AreataDocument10 pagesMethotrexate Alopecia AreataThorn MailNo ratings yet
- Methylene Blue in Anticancer Photodynamic TherapyDocument7 pagesMethylene Blue in Anticancer Photodynamic Therapyenriquecamacho.buyNo ratings yet
- Paper TranscripcionDocument13 pagesPaper TranscripcionKarime MontenegroNo ratings yet
- Kanji 2018Document6 pagesKanji 2018KEVIN ROGGER OBANDO BALLARDONo ratings yet
- Loss-Of-Function Genetic Tools For Animal Models - Cross-Species and Cross-Platform DifferencesDocument17 pagesLoss-Of-Function Genetic Tools For Animal Models - Cross-Species and Cross-Platform DifferencesLeon PalomeraNo ratings yet
- A Review of Methods To Determine Viability, Vitality, and Metabolic Rates in MicrobiologyDocument25 pagesA Review of Methods To Determine Viability, Vitality, and Metabolic Rates in MicrobiologyAna Carolina FurianNo ratings yet
- Paper HydrogelDocument17 pagesPaper HydrogelAntar GayenNo ratings yet
- 2016 Medical Ozone Increases Methotrexate Clinical Response and Improves Cellular Redox Balance in Patients With Rheumatoid ArthritisDocument6 pages2016 Medical Ozone Increases Methotrexate Clinical Response and Improves Cellular Redox Balance in Patients With Rheumatoid ArthritisAcupuntura de EquinosNo ratings yet
- JGM 2990Document7 pagesJGM 2990FaMe TanapornNo ratings yet
- Metabolite Safety in Drug DevelopmentFrom EverandMetabolite Safety in Drug DevelopmentSuzanne L. IversonNo ratings yet
- JGM 2990Document7 pagesJGM 2990FaMe TanapornNo ratings yet
- Pharmacokinetics and Pharmacogenetics of High Dose Methotrexate in Chinese Adult Patients With Non Hodgkin Lymphoma: A Population AnalysisDocument17 pagesPharmacokinetics and Pharmacogenetics of High Dose Methotrexate in Chinese Adult Patients With Non Hodgkin Lymphoma: A Population AnalysisFaMe TanapornNo ratings yet
- Key 275Document4 pagesKey 275FaMe TanapornNo ratings yet
- MTHFR Gene Polymorphisms and Susceptibility To Rheumatoid Arthritis: A Meta-Analysis Based On 16 StudiesDocument13 pagesMTHFR Gene Polymorphisms and Susceptibility To Rheumatoid Arthritis: A Meta-Analysis Based On 16 StudiesFaMe TanapornNo ratings yet
- Association Between MTHFR C677T Polymorphism 2017 PDFDocument11 pagesAssociation Between MTHFR C677T Polymorphism 2017 PDFFaMe TanapornNo ratings yet
- 1070 Impact of MTHFR rs1801133 MTHFR rs1801131 and Abcb1 rs1045642 Polymorphisms With Increased Susceptibility of Rheumatoid Arthritis in The West Algerian Population A Case Control Study FileDocument9 pages1070 Impact of MTHFR rs1801133 MTHFR rs1801131 and Abcb1 rs1045642 Polymorphisms With Increased Susceptibility of Rheumatoid Arthritis in The West Algerian Population A Case Control Study FileFaMe TanapornNo ratings yet
- Acr 23164Document11 pagesAcr 23164FaMe TanapornNo ratings yet
- Z37 PDFDocument8 pagesZ37 PDFFaMe TanapornNo ratings yet
- Regular ArticleDocument8 pagesRegular ArticleFaMe TanapornNo ratings yet
- Use of MTHFR C677T Polymorphism and Plasma Pharmacokinetics To Predict Methotrexate Toxicity in Patients With Acute Lymphoblastic LeukemiaDocument8 pagesUse of MTHFR C677T Polymorphism and Plasma Pharmacokinetics To Predict Methotrexate Toxicity in Patients With Acute Lymphoblastic LeukemiaFaMe TanapornNo ratings yet
- The Journal of Rheumatology Volume 44, No. 6Document8 pagesThe Journal of Rheumatology Volume 44, No. 6FaMe TanapornNo ratings yet
- Sciencedirect: Original ArticleDocument8 pagesSciencedirect: Original ArticleFaMe TanapornNo ratings yet
- Document Analysis Worksheet: Analyze A PaintingDocument4 pagesDocument Analysis Worksheet: Analyze A PaintingPatrick SanchezNo ratings yet
- Swapnil CromaDocument15 pagesSwapnil Cromapanpatil swapnilNo ratings yet
- Medicinal Benefits of HoneyDocument8 pagesMedicinal Benefits of HoneyTauseef AshrafNo ratings yet
- Zabala Auto SupplyDocument7 pagesZabala Auto SupplyLeighsen VillacortaNo ratings yet
- What Is The Difference Between Graduate and UndergraduateDocument6 pagesWhat Is The Difference Between Graduate and UndergraduateWinnie OchiengNo ratings yet
- CME Applique TutorialDocument20 pagesCME Applique TutorialMarittaKarmaNo ratings yet
- (1899) (Vol. 2) A Sailor's Life Under Four SovereignsDocument364 pages(1899) (Vol. 2) A Sailor's Life Under Four SovereignsHerbert Hillary Booker 2ndNo ratings yet
- ToninaDocument6 pagesToninapancholopez2014No ratings yet
- HZ2322 EM250 Series 100mW ZigBee Module (ENG)Document10 pagesHZ2322 EM250 Series 100mW ZigBee Module (ENG)MicroComSolutionsNo ratings yet
- Artigo Hambate BaDocument19 pagesArtigo Hambate BaRayza GiardiniNo ratings yet
- DAS AppFormDocument3 pagesDAS AppFormZia HashmiNo ratings yet
- Integrated NLP Train The Trainer BootcampDocument8 pagesIntegrated NLP Train The Trainer BootcampHitesh ParmarNo ratings yet
- Santiago v. Gonzalez PDFDocument2 pagesSantiago v. Gonzalez PDFGervin ArquizalNo ratings yet
- MSDS Abate - 1 - GR-ID-SDS - 02 - GHSDocument34 pagesMSDS Abate - 1 - GR-ID-SDS - 02 - GHSpurwadiNo ratings yet
- 22TCN263 2000 Tieu Chuan Khao Sat Duong o ToDocument109 pages22TCN263 2000 Tieu Chuan Khao Sat Duong o ToTu PhamNo ratings yet
- Vampires Werewolves and Witches Oh MyDocument51 pagesVampires Werewolves and Witches Oh Myangelg29850% (1)
- 2.2 - Concrete Cracked Elastic Stresses StageDocument12 pages2.2 - Concrete Cracked Elastic Stresses StageMarcelo AbreraNo ratings yet
- Guidelines On Good Research PracticeDocument9 pagesGuidelines On Good Research PracticeAndre SetiawanNo ratings yet
- Weaning Meal Plan 7 9 MonthsDocument1 pageWeaning Meal Plan 7 9 MonthsDeborah EvangelineNo ratings yet
- Dark Souls 3 Lore - Father Ariandel & Sister Friede - VaatividyaDocument5 pagesDark Souls 3 Lore - Father Ariandel & Sister Friede - VaatividyashaunNo ratings yet
- PKM Full C.V. Sept 2021Document11 pagesPKM Full C.V. Sept 2021aopera87No ratings yet
- Startup Law OverviewDocument11 pagesStartup Law OverviewAgibise AkanliseNo ratings yet
- VollyballDocument4 pagesVollyballCristina LataganNo ratings yet
- Final ExamDocument3 pagesFinal ExamFaidah PangandamanNo ratings yet
- Unit 5: Implications of Developmental Biology: Teratogenesis: Types and Teratogenic AgentsDocument12 pagesUnit 5: Implications of Developmental Biology: Teratogenesis: Types and Teratogenic AgentsAmar Kant JhaNo ratings yet
- Just Say No To Big Brother's Smart Meters: The Latest in Bio-Hazard Technology by Orlean KoehleDocument148 pagesJust Say No To Big Brother's Smart Meters: The Latest in Bio-Hazard Technology by Orlean KoehleVen GeanciaNo ratings yet
- Diagnosis of Stroke-Associated PneumoniaDocument16 pagesDiagnosis of Stroke-Associated PneumoniaCecilia Casandra UneputtyNo ratings yet