Download as pdf or txt
You might also like
- Chapter 35Document9 pagesChapter 35Jyothis James100% (2)
- 24 Butterflies Patricia GraceDocument8 pages24 Butterflies Patricia Gracemisspau010No ratings yet
- Broadleaf ForestsDocument10 pagesBroadleaf ForestsScribdTranslationsNo ratings yet
- Tropical SubhumidDocument5 pagesTropical SubhumidScribdTranslationsNo ratings yet
- Forest Types of PakistanDocument13 pagesForest Types of Pakistanbutterfily80% (46)
- Components: Forests) Are More Species-Rich Than Those DominatedDocument6 pagesComponents: Forests) Are More Species-Rich Than Those DominatedLeoNo ratings yet
- FORESTDocument26 pagesFORESTrhose mama100% (1)
- Descriptions of The 16 Major Forest Type-Groups According To Champion and Seth (1968)Document4 pagesDescriptions of The 16 Major Forest Type-Groups According To Champion and Seth (1968)RajKumarNo ratings yet
- Ecosystem of Mexico Gulf: FYBCOM-D-049-SONAM-UPADHYAY-47Document21 pagesEcosystem of Mexico Gulf: FYBCOM-D-049-SONAM-UPADHYAY-47sonam upadhyayNo ratings yet
- Forest TypesDocument10 pagesForest TypesMehboob AlamNo ratings yet
- Forests of PakistanDocument12 pagesForests of PakistanMuhammad Waleed ChaudharyNo ratings yet
- Forest Types of Pak BY Muhammadnadeem Student of University of Swat Center For Agriculture Sciences and ForestryDocument11 pagesForest Types of Pak BY Muhammadnadeem Student of University of Swat Center For Agriculture Sciences and ForestryMuhammad Nadeem100% (1)
- Lesson 4 Philippine Forest and Wildlife Resources: By: For. Leslie Sanchez Obiso CTU-BariliDocument12 pagesLesson 4 Philippine Forest and Wildlife Resources: By: For. Leslie Sanchez Obiso CTU-BariliLeslie ObisoNo ratings yet
- Types of RainforestsDocument5 pagesTypes of RainforestsLiliana TiuNo ratings yet
- Makalah Tropical Rain ForestDocument14 pagesMakalah Tropical Rain Forestgentong_cantikNo ratings yet
- Pacific Tropical ForestDocument17 pagesPacific Tropical ForestScribdTranslationsNo ratings yet
- University of Gujrat: Environmental SciencesDocument11 pagesUniversity of Gujrat: Environmental SciencesSalmanNaseerNo ratings yet
- Forest Types of PakistanDocument6 pagesForest Types of PakistanAryaan GeeNo ratings yet
- Grassland GrasslandDocument9 pagesGrassland GrasslandMahipal SinghNo ratings yet
- Lesson 1 BiomesDocument14 pagesLesson 1 BiomesJim Roger Malabo LorenzoNo ratings yet
- Types of ForestsDocument5 pagesTypes of ForestsNamitNo ratings yet
- Biomes: Global Patterns of Life: By: Felipe C. MarquesDocument22 pagesBiomes: Global Patterns of Life: By: Felipe C. MarquesFelipe Cury MarquesNo ratings yet
- Some Relief Features of East AfricaDocument2 pagesSome Relief Features of East AfricaJoseph ZotooNo ratings yet
- 7 Forest and WildlifeDocument32 pages7 Forest and WildlifeFrances Joei O. PEREZNo ratings yet
- Forests Account For 75Document4 pagesForests Account For 75GrayNo ratings yet
- Types of Forest Ecosystems 2Document13 pagesTypes of Forest Ecosystems 2Karthick Ramamoorthy100% (1)
- Forest ManagementDocument27 pagesForest ManagementAlistair HernandoNo ratings yet
- BiomesDocument3 pagesBiomesJESMELYN RAMOSNo ratings yet
- Tundra Is The Coldest of All The BiomesDocument24 pagesTundra Is The Coldest of All The BiomesAira GarciaNo ratings yet
- A Multiple-Site Comparison of Woodpecker Communities in Bornean Lowland and Hill ForestsDocument12 pagesA Multiple-Site Comparison of Woodpecker Communities in Bornean Lowland and Hill ForestsChitoge GremoryNo ratings yet
- Social (Geography) Group ProjectDocument17 pagesSocial (Geography) Group ProjectlittlevishnuckNo ratings yet
- Mediterranean Forests, Woodlands, and ScrubDocument5 pagesMediterranean Forests, Woodlands, and Scrubsorbciren13No ratings yet
- Sci 2Document6 pagesSci 2Jvyn GamingNo ratings yet
- Proposa LDocument19 pagesProposa LLea Joy CagbabanuaNo ratings yet
- Natural Vegetation and Wildlife: Made by Prashant KumarDocument18 pagesNatural Vegetation and Wildlife: Made by Prashant KumarLegendNo ratings yet
- Forest-types-of-Pak-with-sub-division by Muhammad Nadeem Student of MPhil Forestry University of SwatDocument11 pagesForest-types-of-Pak-with-sub-division by Muhammad Nadeem Student of MPhil Forestry University of Swatnadeem khanNo ratings yet
- DeforestationDocument7 pagesDeforestationAyasha MalagaNo ratings yet
- ENGG 413 Environmental Science and Engineering Major Ecosystem of The WorldDocument20 pagesENGG 413 Environmental Science and Engineering Major Ecosystem of The WorldFatima Therese ManaloNo ratings yet
- Serra Do Mar Coastal ForestsDocument3 pagesSerra Do Mar Coastal Forestssamy.s.extraNo ratings yet
- BiomesDocument9 pagesBiomesapi-26570979100% (1)
- Forest and Wildlife: Made By:-Dipit Singh Kanwar Academic Year:-2020/21 Teacher: - Amrita JoganiDocument18 pagesForest and Wildlife: Made By:-Dipit Singh Kanwar Academic Year:-2020/21 Teacher: - Amrita Joganivineet_knwrNo ratings yet
- Major Biomes of The WorldDocument4 pagesMajor Biomes of The WorldNaga Chary100% (1)
- GE15 ULObDocument7 pagesGE15 ULObKyle Adrian MartinezNo ratings yet
- Imp of ForestDocument8 pagesImp of ForestShikha GuptaNo ratings yet
- Field Study Tour - Yamuna Biodiversity Park - Brocvhure and Instructions - Format For ReportDocument8 pagesField Study Tour - Yamuna Biodiversity Park - Brocvhure and Instructions - Format For Reportaditya doneriaNo ratings yet
- Denr DMC 2005-005Document5 pagesDenr DMC 2005-005DavidNo ratings yet
- Natural Ecosystems in The PhilippinesDocument3 pagesNatural Ecosystems in The PhilippinesJelian ReyesNo ratings yet
- Layers: Trees Shrubs Vines Grasses Mosses Algae Fungi Insects Mammals Birds Reptiles Amphibians EditDocument3 pagesLayers: Trees Shrubs Vines Grasses Mosses Algae Fungi Insects Mammals Birds Reptiles Amphibians EditZoe FallurinNo ratings yet
- FORESTRYDocument10 pagesFORESTRYkwambokasly21No ratings yet
- Forest and Philippine Wildlife and ResourcesDocument54 pagesForest and Philippine Wildlife and ResourcesMarian Alexis FernandezNo ratings yet
- Tundra Taiga Temperate Deciduous Forest Scrub Forest Grassland Desert Tropical Rain Forest Temperate Rain ForestDocument11 pagesTundra Taiga Temperate Deciduous Forest Scrub Forest Grassland Desert Tropical Rain Forest Temperate Rain Forestvenus caalemNo ratings yet
- Global Geography 14Document6 pagesGlobal Geography 14IliasNo ratings yet
- Biology: Specific ObjectivesDocument8 pagesBiology: Specific ObjectivesCarlos Felino LazatinNo ratings yet
- Rainforest Booklet PDFDocument20 pagesRainforest Booklet PDFdefiaaaNo ratings yet
- Biomes of Our EarthDocument5 pagesBiomes of Our EarthMayaNo ratings yet
- Natural Resources Management: RMGT 230Document41 pagesNatural Resources Management: RMGT 230GRACE VERIDIANONo ratings yet
- Poster Forests December 11 Inside 172-173Document1 pagePoster Forests December 11 Inside 172-173veldfloraedNo ratings yet
- Sundaland Heath Forests - Ecoregions - WWFDocument4 pagesSundaland Heath Forests - Ecoregions - WWFErna Karlinna D. YanthyNo ratings yet
- Tropical ForestsDocument8 pagesTropical Forestsadi govindNo ratings yet
- All About RainforestDocument4 pagesAll About Rainforestapi-266141300No ratings yet
- The Complete U.S. Army Survival Guide to Tropical, Desert, Cold Weather, Mountain Terrain, Sea, and NBC EnvironmentsFrom EverandThe Complete U.S. Army Survival Guide to Tropical, Desert, Cold Weather, Mountain Terrain, Sea, and NBC EnvironmentsNo ratings yet
- Seed GerminationDocument12 pagesSeed Germinationhrishikesh08No ratings yet
- Bitter-Orange-epicarp and Mesocarp TinctureDocument2 pagesBitter-Orange-epicarp and Mesocarp TinctureNikotiano TobacumNo ratings yet
- Presenting Insights and Findings: Written Reports: CHAPTER 19 - 20Document20 pagesPresenting Insights and Findings: Written Reports: CHAPTER 19 - 20Eno CasmiNo ratings yet
- Bryophytes of TNDocument117 pagesBryophytes of TNHabib UllahNo ratings yet
- Attic24 - May Roses CrochetDocument15 pagesAttic24 - May Roses CrochetAnonymous FXsb2MNo ratings yet
- SesameDocument6 pagesSesameSuhara BeevyNo ratings yet
- Modul Perfect Score SBP Biology SPM 2013 Question and SchemeDocument283 pagesModul Perfect Score SBP Biology SPM 2013 Question and SchemeCikgu Faizal87% (15)
- Bamboo Plant Pests and DiseasesDocument4 pagesBamboo Plant Pests and DiseasesSynkroMax BiotechNo ratings yet
- Annies Calla LilliesDocument8 pagesAnnies Calla Lilliesbuffylovez100% (2)
- MarchantiophytaDocument9 pagesMarchantiophytax456456456xNo ratings yet
- MCQ ch2Document4 pagesMCQ ch2Anirudh CNo ratings yet
- Botanical Monograph Project BelladonnaDocument7 pagesBotanical Monograph Project BelladonnaAbdullah A. ElgazarNo ratings yet
- Keys To The Genera of Annonaceae.xDocument10 pagesKeys To The Genera of Annonaceae.xMaikol GuevaraNo ratings yet
- The Local Knowledge of Medicinal Plants Trader and Diversity of Medicinal Plants in The Kabanjahe Traditional Market, North Sumatra, IndonesiaDocument12 pagesThe Local Knowledge of Medicinal Plants Trader and Diversity of Medicinal Plants in The Kabanjahe Traditional Market, North Sumatra, IndonesialenteracintaNo ratings yet
- CarrotsDocument3 pagesCarrotsemaNo ratings yet
- Tannin From Betel Nut (Areca Catechu) As An Alternative Dewormer For Broiler Chickens (Gallus Gallus Domesticus)Document10 pagesTannin From Betel Nut (Areca Catechu) As An Alternative Dewormer For Broiler Chickens (Gallus Gallus Domesticus)Madrick Ace VillaNo ratings yet
- Amphibia Anne or Beast Storyboard First SceneDocument339 pagesAmphibia Anne or Beast Storyboard First ScenenanopulgaNo ratings yet
- Indiana Rain Garden Plant List Recommended For Jeffersonville - City of JeffersonvilleDocument3 pagesIndiana Rain Garden Plant List Recommended For Jeffersonville - City of JeffersonvilleFree Rain Garden Manuals and MoreNo ratings yet
- What Is Capiz ShellDocument4 pagesWhat Is Capiz ShellIra James AbanesNo ratings yet
- Lesson 1.2 - The Mathematics in Our WorldDocument52 pagesLesson 1.2 - The Mathematics in Our WorldGuia CandelariaNo ratings yet
- Rules 2019 Chapter 0Document14 pagesRules 2019 Chapter 0Frederico DellanoNo ratings yet
- M.sc. Botany2010Document24 pagesM.sc. Botany2010Vishal AgrahariNo ratings yet
- Plant Tissues & AnatomyDocument25 pagesPlant Tissues & Anatomyoption 360No ratings yet
- Life Mastery Eating Plan PDFDocument22 pagesLife Mastery Eating Plan PDFCatalinNo ratings yet
- Adenium Obesum Hybrid Pink PDFDocument3 pagesAdenium Obesum Hybrid Pink PDFRohit PatilNo ratings yet
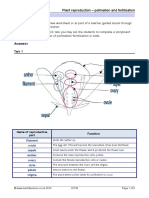
- Pollination and FertilisationDocument5 pagesPollination and FertilisationmihayedoNo ratings yet
- Biology Plant TissuesDocument10 pagesBiology Plant TissuesChandrakala K RNo ratings yet
- Cet Biology: 1 and 2 PUC Biology PracticalsDocument44 pagesCet Biology: 1 and 2 PUC Biology PracticalsZaina RehmanNo ratings yet