Download as pdf or txt
You might also like
- BGAS Theory PaperDocument22 pagesBGAS Theory PaperAdilMunir100% (3)
- Unit 1: Laboratory ManualDocument60 pagesUnit 1: Laboratory ManualPatrice HomerNo ratings yet
- Hydrosulphite DeterminationDocument3 pagesHydrosulphite Determination950 911100% (1)
- Heavy Metal Resistance in Transgenic PlantsDocument6 pagesHeavy Metal Resistance in Transgenic PlantsAlexandra-DianaAlexandraNo ratings yet
- Heavy Metal Stress and Crop Productivity: December 2015Document26 pagesHeavy Metal Stress and Crop Productivity: December 2015Simran AroraNo ratings yet
- Is Lithium Biologically An Important or Toxic ElemDocument14 pagesIs Lithium Biologically An Important or Toxic ElemLtNo ratings yet
- Mutation Research/Genetic Toxicology and Environmental MutagenesisDocument7 pagesMutation Research/Genetic Toxicology and Environmental MutagenesisrachiiidaNo ratings yet
- Literature Review of Heavy Metals in SoilDocument6 pagesLiterature Review of Heavy Metals in Soilaflskdwol100% (1)
- Government College University Faisalabad: Department of BotanyDocument17 pagesGovernment College University Faisalabad: Department of BotanySaba AnjumNo ratings yet
- Analysis of Antioxidant Enzyme Activity During Germination of Alfalfa Under Salt and Drought StressesDocument8 pagesAnalysis of Antioxidant Enzyme Activity During Germination of Alfalfa Under Salt and Drought StressesJimenoNo ratings yet
- Phytotoxicity and Phytoremediation Potential of Mercury in Indian Mustard and Two Ferns With Mercury Contaminated Water and Oak Ridge Soil - 9241Document12 pagesPhytotoxicity and Phytoremediation Potential of Mercury in Indian Mustard and Two Ferns With Mercury Contaminated Water and Oak Ridge Soil - 9241レテイ サマ ヒルカNo ratings yet
- The Growth and Antioxidative Responses of Cu (II), PB (II), CD (II) and CR (VI) Stress ConditionDocument13 pagesThe Growth and Antioxidative Responses of Cu (II), PB (II), CD (II) and CR (VI) Stress ConditionAli Zainal Enal ShahabNo ratings yet
- Phytoremediation of Silver Nano ParticlesDocument7 pagesPhytoremediation of Silver Nano Particlesaltair001No ratings yet
- FPBGriselda 2016Document13 pagesFPBGriselda 2016Griselda PodazzaNo ratings yet
- CR 5 PDFDocument11 pagesCR 5 PDFام احمدNo ratings yet
- KhoshkholghSimaetal 2013CoparativeEffectsofSaltsDocument19 pagesKhoshkholghSimaetal 2013CoparativeEffectsofSaltsØp Lone WølfNo ratings yet
- Heavymetals 2Document11 pagesHeavymetals 2hilmiNo ratings yet
- Agronomy 11 00131Document18 pagesAgronomy 11 00131Jose LuisNo ratings yet
- Plants: Jute: A Potential Candidate For Phytoremediation of Metals-A ReviewDocument14 pagesPlants: Jute: A Potential Candidate For Phytoremediation of Metals-A ReviewDark KillerNo ratings yet
- Verma 2003Document11 pagesVerma 2003gdriveanas gdriveNo ratings yet
- Antioxidants y Antioxidant EnzymesDocument306 pagesAntioxidants y Antioxidant EnzymesDTipantaNo ratings yet
- Myco Chemtech PDFDocument8 pagesMyco Chemtech PDFPepy F NataayuNo ratings yet
- Laccase EnzymeDocument12 pagesLaccase EnzymeAnwar DanishNo ratings yet
- Indu MamDocument13 pagesIndu MamRohit AroraNo ratings yet
- Cadmio BiorremediacionDocument21 pagesCadmio BiorremediacionDANIA ALEJANDRA POVEDA MURCIANo ratings yet
- Tagetes Erecta (Marigold), A Phytoremediant For Ni-And Pb-Contaminated Area: A Hydroponic Analysis and Factors InvolvedDocument7 pagesTagetes Erecta (Marigold), A Phytoremediant For Ni-And Pb-Contaminated Area: A Hydroponic Analysis and Factors InvolvedMaria AnggieNo ratings yet
- The Potential of Selenium To Induce Salt Stress ToleranceDocument14 pagesThe Potential of Selenium To Induce Salt Stress Tolerancewaheed akramNo ratings yet
- 1 s2.0 S0045653522005380 Main PDFDocument12 pages1 s2.0 S0045653522005380 Main PDFmishell baltazarNo ratings yet
- Isolation, Characterization and Biosorption of Zinc by Indigenous Fungal Strains Aspergillus Fumigatus RH05 and Aspergillus Avus RH07Document17 pagesIsolation, Characterization and Biosorption of Zinc by Indigenous Fungal Strains Aspergillus Fumigatus RH05 and Aspergillus Avus RH07Wiliam CasasNo ratings yet
- Chandra - 2017 - Phytoextraction of Heavy Metals by Potential Native PlantsDocument15 pagesChandra - 2017 - Phytoextraction of Heavy Metals by Potential Native Plantsiki lunisaNo ratings yet
- RiceDocument14 pagesRiceMD. HasanNo ratings yet
- Citric Acido Plants-08-00525Document16 pagesCitric Acido Plants-08-00525Sergio Luís Fernandes PereiraNo ratings yet
- Roleofmushroomsinsoilmycoremediation AreviewDocument10 pagesRoleofmushroomsinsoilmycoremediation AreviewS6rNo ratings yet
- Concentration of Heavy Metals in Root, Stem and LeavesDocument9 pagesConcentration of Heavy Metals in Root, Stem and LeavesladipupoNo ratings yet
- Afshan Et Al 2015 CA On CR On Brassica ESPRDocument11 pagesAfshan Et Al 2015 CA On CR On Brassica ESPREyeNo ratings yet
- Dnapuri Ref5 PDFDocument8 pagesDnapuri Ref5 PDFHumaNazNo ratings yet
- Evaluation and Treatment of PB, CD, and Ni by Using Four Water Plants Belonging To (Ceratophyllum SPP.) For Wastewater in Tikrit Teaching HospitalDocument16 pagesEvaluation and Treatment of PB, CD, and Ni by Using Four Water Plants Belonging To (Ceratophyllum SPP.) For Wastewater in Tikrit Teaching HospitalCentral Asian StudiesNo ratings yet
- 1,2 JugaDocument7 pages1,2 JugaDita MarthariniNo ratings yet
- Journal of Stress Physiology & BiochemistryDocument9 pagesJournal of Stress Physiology & BiochemistryИмямямя ФамилилилилияNo ratings yet
- Research Paper Published On Bioremediation of Heavy Metals From AlgaeDocument9 pagesResearch Paper Published On Bioremediation of Heavy Metals From AlgaeaflbtvmfpNo ratings yet
- Bankaji2014 Article EffectsOfCombinedAbioticStressDocument12 pagesBankaji2014 Article EffectsOfCombinedAbioticStressLydia CasasniNo ratings yet
- Cadmium Toxicity Symptoms and Uptake Mechanism in PlantsDocument17 pagesCadmium Toxicity Symptoms and Uptake Mechanism in PlantsAlineNo ratings yet
- Phyto 1Document12 pagesPhyto 1MA RI AHNo ratings yet
- Antioxidative and Biochemical Responses in Dalberg PDFDocument7 pagesAntioxidative and Biochemical Responses in Dalberg PDFmannjaviNo ratings yet
- Response of Key Soil Parameters During Compost-Assisted Phytostabilization in Extremely Acidic Tailings: Effect of Plant SpeciesDocument9 pagesResponse of Key Soil Parameters During Compost-Assisted Phytostabilization in Extremely Acidic Tailings: Effect of Plant Speciesgor45No ratings yet
- Oxidative Stress Reactive Oxygen Species Regulation and ROS Scavenging Pathways in PlantsDocument6 pagesOxidative Stress Reactive Oxygen Species Regulation and ROS Scavenging Pathways in PlantsChhan Kumar kalitaNo ratings yet
- Drug Invention TodayDocument7 pagesDrug Invention TodaySavariraj Rajkumar DominicNo ratings yet
- Fitoestracción y RizosecreciónDocument5 pagesFitoestracción y RizosecreciónMane Becerril MerlosNo ratings yet
- Plant Physiology and Biochemistry: Houssem Kahli, Hana Sbartai, Touria Cohen-Bouhacina, Jacques BourguignonDocument10 pagesPlant Physiology and Biochemistry: Houssem Kahli, Hana Sbartai, Touria Cohen-Bouhacina, Jacques BourguignonAmina HdNo ratings yet
- tmpB9EB TMPDocument16 pagestmpB9EB TMPFrontiersNo ratings yet
- Comparison of Seed Germination and Recovery Responses of A Salt Marsh Halophyte Halopeplis Perfoliata To Osmotic and Ionic TreatmentsDocument10 pagesComparison of Seed Germination and Recovery Responses of A Salt Marsh Halophyte Halopeplis Perfoliata To Osmotic and Ionic TreatmentsAsaad LahmarNo ratings yet
- CD Phytotoxicity Issues, Progress, Enviromental Concerns and Future PerspectivesDocument15 pagesCD Phytotoxicity Issues, Progress, Enviromental Concerns and Future PerspectivesKarina LizethNo ratings yet
- Sairam 2019Document8 pagesSairam 2019Malik HopeNo ratings yet
- Lead and Cadmium Uptake in The Marine Fungi Corollospora Lacera and Monodictys PelagicaDocument9 pagesLead and Cadmium Uptake in The Marine Fungi Corollospora Lacera and Monodictys PelagicaLyca Khate AbanNo ratings yet
- Biochemical and Functional Responses of Arabidopsis Thaliana Exposed To Cadmium, Copper and ZincDocument25 pagesBiochemical and Functional Responses of Arabidopsis Thaliana Exposed To Cadmium, Copper and ZincAprilia azzahraNo ratings yet
- De Novo Transcriptome and Small RNA Analysis of Two Chinese Willow Cultivars Reveals Stress Response Genes in Salix MatsudanaDocument12 pagesDe Novo Transcriptome and Small RNA Analysis of Two Chinese Willow Cultivars Reveals Stress Response Genes in Salix MatsudanaHogarth FrankfurtNo ratings yet
- Literature Review On Heavy Metals in VegetablesDocument8 pagesLiterature Review On Heavy Metals in Vegetablesbij0dizytaj2100% (1)
- Changes of Organic Acid Exudation and Rhizosphere PH in Rice Plants Under Chromium StressDocument6 pagesChanges of Organic Acid Exudation and Rhizosphere PH in Rice Plants Under Chromium StressJonnatan PelawzNo ratings yet
- Plant Growth Promoting Curtobacterium Albidum Strain SRV4: An Agriculturally Important Microbe To Alleviate Salinity Stress in Paddy PlantsDocument11 pagesPlant Growth Promoting Curtobacterium Albidum Strain SRV4: An Agriculturally Important Microbe To Alleviate Salinity Stress in Paddy PlantsmileidyNo ratings yet
- Abiotic Stress Alleviation With Brassinosteroids in Plant RootsDocument22 pagesAbiotic Stress Alleviation With Brassinosteroids in Plant RootsArya ChowdhuryNo ratings yet
- Plants 12 03792Document21 pagesPlants 12 03792Bqshalihatun PutriNo ratings yet
- Modern Rhodium-Catalyzed Organic ReactionsFrom EverandModern Rhodium-Catalyzed Organic ReactionsP. Andrew EvansNo ratings yet
- Reactive Oxygen Species in Plants: Boon Or Bane - Revisiting the Role of ROSFrom EverandReactive Oxygen Species in Plants: Boon Or Bane - Revisiting the Role of ROSDr. Vijay Pratap SinghNo ratings yet
- Candle Holder ReportDocument7 pagesCandle Holder ReportSHARIFAH AZIAH SYED RIDHUANNo ratings yet
- Crude OilDocument2 pagesCrude OilBasia JakubiakNo ratings yet
- University of Central Punjab Faculty of SciencesDocument18 pagesUniversity of Central Punjab Faculty of SciencesAttitudekiller RanaNo ratings yet
- Gelling Agent: Ips Academy College of Pharmacy IndoreDocument42 pagesGelling Agent: Ips Academy College of Pharmacy Indoreayushi shuklaNo ratings yet
- Chapter I Composition of Matter Marilou ELBODocument5 pagesChapter I Composition of Matter Marilou ELBOTn F'dzNo ratings yet
- CV 1Document11 pagesCV 1Misaki 1No ratings yet
- Indiana University Doctoral DissertationsDocument4 pagesIndiana University Doctoral DissertationsFindSomeoneToWriteMyPaperLittleRock100% (1)
- HW - Carbohydrate Metabolism II & Lipid MetabolismDocument2 pagesHW - Carbohydrate Metabolism II & Lipid MetabolismyanNo ratings yet
- Tetrahydro Quino LinesDocument40 pagesTetrahydro Quino LinesРумен ЛяпчевNo ratings yet
- Class - XII (Frequntly Asked Topics)Document13 pagesClass - XII (Frequntly Asked Topics)yrh6qgd2xnNo ratings yet
- 30 MW Power PlantDocument151 pages30 MW Power Plantsambhu0% (1)
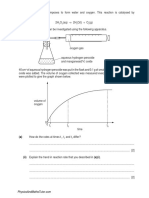
- 1 Hydrogen Peroxide Decomposes To Form Water and Oxygen. This Reaction Is Catalysed byDocument9 pages1 Hydrogen Peroxide Decomposes To Form Water and Oxygen. This Reaction Is Catalysed byKelvin DenhereNo ratings yet
- The Learners Demonstrate Understandin G Of... The Learners Should Be Able To..Document8 pagesThe Learners Demonstrate Understandin G Of... The Learners Should Be Able To..Bryan BejeranoNo ratings yet
- Construction Biotechnology: Volodymyr Ivanov Viktor StabnikovDocument332 pagesConstruction Biotechnology: Volodymyr Ivanov Viktor StabnikovHector Santiago Lopez AcostaNo ratings yet
- Watson and Crick Model of DNADocument22 pagesWatson and Crick Model of DNAGanesh V Gaonkar100% (1)
- JHANVI SAREEN - Biochem Assignment (2.0)Document9 pagesJHANVI SAREEN - Biochem Assignment (2.0)Jhanvi sareenNo ratings yet
- Ha Coating On Ti 3.1Document9 pagesHa Coating On Ti 3.1Truong DucQuangNo ratings yet
- Comprehensive Organic Synthesis Volume 1 1991Document987 pagesComprehensive Organic Synthesis Volume 1 1991Paul YoungNo ratings yet
- Rates of Reaction Chapter 8Document2 pagesRates of Reaction Chapter 8Mariam BabarNo ratings yet
- 4.06 - 4.07 Aromatic Chemistry and Amines MSDocument20 pages4.06 - 4.07 Aromatic Chemistry and Amines MSAdnan ChowdhuryNo ratings yet
- Production of Drying OilDocument136 pagesProduction of Drying Oilali abdulrahman al-ezziNo ratings yet
- Benecel™ Methylcellulose, Hypromellose and Methylhydroxyethyl CelluloseDocument3 pagesBenecel™ Methylcellulose, Hypromellose and Methylhydroxyethyl CelluloseJose Andres RodriguezNo ratings yet
- ChromatographyDocument40 pagesChromatographydarshanNo ratings yet
- Inverted Sugar Syrup Attained From Sucrose HydrolyDocument8 pagesInverted Sugar Syrup Attained From Sucrose HydrolySiti AmirahNo ratings yet
- Bspil 573 S Photosynthesis LightDocument1 pageBspil 573 S Photosynthesis LightJagdish RajanNo ratings yet
- Land Pollution and SWMDocument126 pagesLand Pollution and SWMShawn BaynoNo ratings yet
- Module 3C - Cellular RespirationDocument9 pagesModule 3C - Cellular RespirationDiane Balaba OsingNo ratings yet