
Simple Lipids:: Characteristics of Each Are Described in The Sections Below
Simple Lipids:: Characteristics of Each Are Described in The Sections Below
You might also like
- Stoker Chapter19 (Lipids) - 2 PDFDocument85 pagesStoker Chapter19 (Lipids) - 2 PDFAsther Mantua86% (7)
- Experiment 5 LipidsDocument13 pagesExperiment 5 LipidsClemence Marie FuentesNo ratings yet
- 08LipAMO SuarezDocument15 pages08LipAMO SuarezscasuarezNo ratings yet
- Lipid MetabolismDocument32 pagesLipid MetabolismNahid HasanNo ratings yet
- Chapter 08 LipidDocument10 pagesChapter 08 Lipid楊畯凱No ratings yet
- SESE 115 Biochemistry: Don Mariano Marcos Memorial State University La Union PhilippinesDocument13 pagesSESE 115 Biochemistry: Don Mariano Marcos Memorial State University La Union PhilippinesVhenz MapiliNo ratings yet
- Handout 4 - LipidsDocument7 pagesHandout 4 - LipidsPaul CliffordNo ratings yet
- UntitledDocument7 pagesUntitledGodstime sakaNo ratings yet
- Structure of Membrane LipidsDocument41 pagesStructure of Membrane LipidsStanley ChikoveNo ratings yet
- Unit-Iv-Lipids and PorphyrinsDocument29 pagesUnit-Iv-Lipids and PorphyrinsNila ArivoliNo ratings yet
- 05 - Intro UNIT 4Document23 pages05 - Intro UNIT 4Elena OlmedoNo ratings yet
- Biological MacromoleculesDocument2 pagesBiological MacromoleculesBeyoung GamefightNo ratings yet
- Basic Metabolism II Notes Reviewed-1Document80 pagesBasic Metabolism II Notes Reviewed-1felixNo ratings yet
- Chemistry and Digestion of LipidsDocument35 pagesChemistry and Digestion of LipidsBiph BiphNo ratings yet
- BIO 121 Tutorial 14-07-2021Document19 pagesBIO 121 Tutorial 14-07-2021PARVATHY ANIL - IMS20211No ratings yet
- MC2 Biochemistry Lecture Notes For BSN First Semester, 2019-2020 Prepared By: SALINA OSIAL - ALFADDocument5 pagesMC2 Biochemistry Lecture Notes For BSN First Semester, 2019-2020 Prepared By: SALINA OSIAL - ALFADAl-waleed JulkanainNo ratings yet
- CHEM 140 Unit 8 Lec 1 LipidsDocument93 pagesCHEM 140 Unit 8 Lec 1 LipidsMevil Jane MabrasNo ratings yet
- L3 LipidsDocument38 pagesL3 LipidsAman ImranNo ratings yet
- Biochemistry of Lipids PDFDocument51 pagesBiochemistry of Lipids PDFZayan HaiderNo ratings yet
- Different Types of LipidsDocument7 pagesDifferent Types of LipidsNESLIHAN ERSOYAKNo ratings yet
- Biochem Exam 3 Study GuideDocument17 pagesBiochem Exam 3 Study GuideGreg ZhangNo ratings yet
- (Biochem A) 1.2 Lipid Chemistry (Viliran)Document12 pages(Biochem A) 1.2 Lipid Chemistry (Viliran)CaeNo ratings yet
- Lipids BiochemDocument2 pagesLipids BiochemDoc Zay VillafuerteNo ratings yet
- LIPID (Lec) 2019Document7 pagesLIPID (Lec) 2019Joanna Marie TulioNo ratings yet
- 2.1.2 Biological Molecules Part 2Document7 pages2.1.2 Biological Molecules Part 2sjNo ratings yet
- Biological Molecules and EnzymesDocument16 pagesBiological Molecules and EnzymesdebbyhooiNo ratings yet
- 2 LipidsDocument90 pages2 LipidsHasen umerNo ratings yet
- Biological Molecules 2: Carbohydrates and LipidsDocument53 pagesBiological Molecules 2: Carbohydrates and LipidsKyla Cheyenne BustosNo ratings yet
- Bio MoleculesDocument6 pagesBio Moleculesoption 360No ratings yet
- Lipids 2.sabDocument18 pagesLipids 2.sabShujaNo ratings yet
- Fats, Oils, Phospholipids, Waxes, Steroids, Soaps: BL10A HALL 2006 1Document34 pagesFats, Oils, Phospholipids, Waxes, Steroids, Soaps: BL10A HALL 2006 1cbar biologyNo ratings yet
- Smomo Dani and Luvo Bio PresentationDocument3 pagesSmomo Dani and Luvo Bio PresentationGugu MthethwaNo ratings yet
- 2-Biological MoleculesDocument47 pages2-Biological MoleculesaithanjuNo ratings yet
- Lecture 2 Lipids 3Document23 pagesLecture 2 Lipids 3Caamir Dek HaybeNo ratings yet
- Lipids Are A Diverse Group of Hydrophobic MoleculesDocument5 pagesLipids Are A Diverse Group of Hydrophobic MoleculesDanielle GuerraNo ratings yet
- The Chemical Constituents of Cells1Document14 pagesThe Chemical Constituents of Cells1ArnelBautistaNo ratings yet
- Notes On Lipids and Nucleic Acids-Df 19-1Document21 pagesNotes On Lipids and Nucleic Acids-Df 19-1Albert MweembaNo ratings yet
- C46 Lec6 LipidsDocument91 pagesC46 Lec6 LipidsMary Aniderf Cagape Maturan-GumanoyNo ratings yet
- BiomoleculesDocument4 pagesBiomoleculesSchuyler PeytonNo ratings yet
- Module 3 LipidsDocument8 pagesModule 3 LipidsAiza OrdonoNo ratings yet
- Lipids LehningerDocument7 pagesLipids LehningerElla BangalanNo ratings yet
- 10.2 Lipids IntroductionDocument55 pages10.2 Lipids Introductionperssivesimangavwa2002No ratings yet
- Lecture 2 - Introductory BiochemistryDocument15 pagesLecture 2 - Introductory BiochemistryJana-Tae KerrNo ratings yet
- Structure and Classification of Lipids: Lipids Are Naturally Occurring MoleculesDocument19 pagesStructure and Classification of Lipids: Lipids Are Naturally Occurring MoleculesKomic WebtoonNo ratings yet
- Topic-1A (Food and Health) (Autosaved) - 1-20Document20 pagesTopic-1A (Food and Health) (Autosaved) - 1-20lisaNo ratings yet
- BS1001 LipidsDocument32 pagesBS1001 LipidsLiloyi LupiyaNo ratings yet
- BIOMOLECULES EditedDocument4 pagesBIOMOLECULES Editedtgeorgina630No ratings yet
- Pre-Labs 2 Organic Compositions of The CellDocument6 pagesPre-Labs 2 Organic Compositions of The CellTrần Xuân QuỳnhNo ratings yet
- Fatty Acid MetabolismDocument8 pagesFatty Acid MetabolismJessicaOktavianusNo ratings yet
- (Lipid) : Report AboutDocument8 pages(Lipid) : Report AboutMahamad BakrNo ratings yet
- 1.4A Lipid ChemistryDocument12 pages1.4A Lipid ChemistryBea SamonteNo ratings yet
- Chapter 5: Lipids: 5.1. General Considerations About LipidDocument7 pagesChapter 5: Lipids: 5.1. General Considerations About LipidMD. SAJID GHUFRANNo ratings yet
- Biochemistry: Water Carbohydrates Lipids ProteinsDocument16 pagesBiochemistry: Water Carbohydrates Lipids ProteinsryanNo ratings yet
- Digestion and Absorption of LipidsDocument5 pagesDigestion and Absorption of LipidsAngelo Jude Cobacha100% (1)
- Lipids and Nucleic AcidsDocument20 pagesLipids and Nucleic Acidsengr_rimNo ratings yet
- CHO Chemistry: Functions of CarbohydratesDocument3 pagesCHO Chemistry: Functions of CarbohydratesSaadia AnjumNo ratings yet
- Lecture 2 - Introductory BiochemistryDocument17 pagesLecture 2 - Introductory BiochemistryKevonSingh1No ratings yet
- A Level BiologyDocument522 pagesA Level BiologyJAMESNo ratings yet
- 12 - Analysis of LipidsDocument15 pages12 - Analysis of LipidsLIAN KEI HILADONo ratings yet
- Anatomy and Physiology For Students: A College Level Study Guide for Life Science and Allied Health MajorsFrom EverandAnatomy and Physiology For Students: A College Level Study Guide for Life Science and Allied Health MajorsNo ratings yet
- Biochemistry Applied to the Brewing Processes - Fermentation and the Finished BeerFrom EverandBiochemistry Applied to the Brewing Processes - Fermentation and the Finished BeerRating: 5 out of 5 stars5/5 (1)
- C (Dr/Ãfrfi" P¿Z U) HZFT I) LGJL "8L45F86: Lgit Ol Fy (GL H ZL TFZLBMDocument5 pagesC (Dr/Ãfrfi" P¿Z U) HZFT I) LGJL "8L45F86: Lgit Ol Fy (GL H ZL TFZLBMMukeshsinh GadhaviNo ratings yet
- PN 41 2020Document7 pagesPN 41 2020Mukeshsinh GadhaviNo ratings yet
- MA Scheme ExamDocument6 pagesMA Scheme ExamMukeshsinh GadhaviNo ratings yet
- 54 July 1974 Vol. 2 No. 3: Biochemical EducationDocument2 pages54 July 1974 Vol. 2 No. 3: Biochemical EducationMukeshsinh GadhaviNo ratings yet
- Om Prakash PPT BiologyDocument11 pagesOm Prakash PPT BiologyOm Prakash100% (1)
- Desain Tanpa JudulDocument15 pagesDesain Tanpa JudulAprilia NaomiNo ratings yet
- Fixed Oils and Fats: Group 3Document19 pagesFixed Oils and Fats: Group 3Addis MémñøňNo ratings yet
- Nucleic Acid TransDocument2 pagesNucleic Acid TransJae TyNo ratings yet
- Chapter 1 - Part 1 Introduction Classification of Edible OilDocument60 pagesChapter 1 - Part 1 Introduction Classification of Edible OilFirdaus ZubbirNo ratings yet
- Soal CC StrokeDocument1 pageSoal CC StrokeAngeline BudimanNo ratings yet
- Enzyme WorksheetDocument3 pagesEnzyme Worksheetchinmayee chitturu100% (1)
- Arellano University: Physical ScienceDocument22 pagesArellano University: Physical ScienceMa Angelica MasangcayNo ratings yet
- Preparation and Properties of Cheese FudgeDocument10 pagesPreparation and Properties of Cheese FudgeCM MaryNo ratings yet
- Effects of Multi-Enzyme Supplementation On Nutrient Digestibility in Broiler ChickensDocument6 pagesEffects of Multi-Enzyme Supplementation On Nutrient Digestibility in Broiler ChickensOliver TalipNo ratings yet
- BP303TP MergedDocument10 pagesBP303TP MergedManish ChauhanNo ratings yet
- Pcog Assignment Lipids Olfu PharmacyDocument6 pagesPcog Assignment Lipids Olfu Pharmacyadbiral2226cabNo ratings yet
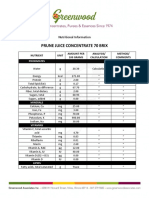
- Prune Juice Concentrate 70 Brix Nutritional InformationDocument2 pagesPrune Juice Concentrate 70 Brix Nutritional Informationborn2dive 9702No ratings yet
- Daksh MahajanDocument12 pagesDaksh MahajanslippypizzaNo ratings yet
- Nonribosomal Peptide Synthetase Biosynthetic Clusters of ESKAPE PathogensDocument29 pagesNonribosomal Peptide Synthetase Biosynthetic Clusters of ESKAPE PathogensMarina DomencoNo ratings yet
- DOPA BiomoleculesDocument9 pagesDOPA Biomoleculesantaryami barikNo ratings yet
- Nucleic AcidsDocument5 pagesNucleic AcidsKheza SuravillaNo ratings yet
- Matdid 207367Document40 pagesMatdid 207367Deblina JanaNo ratings yet
- General Biology 2Document4 pagesGeneral Biology 2Christianne Joseph MonzonNo ratings yet
- Proteins Exam QSDocument5 pagesProteins Exam QSSourav ArunNo ratings yet
- PSmarkup - Jurnal Karya Tulis Ilmiah CiplukanDocument5 pagesPSmarkup - Jurnal Karya Tulis Ilmiah CiplukanrazakNo ratings yet
- Recommended Dietary Allowances - VitaminasDocument3 pagesRecommended Dietary Allowances - VitaminasDaniel Eduardo Lopez CabraNo ratings yet
- animAL Husb. ss2Document1 pageanimAL Husb. ss2Jesse MosesNo ratings yet
- The Central Dogma of Molecular BiologyDocument45 pagesThe Central Dogma of Molecular BiologyEirla Kneriz Trinidad67% (3)
- Heat Shock Proteins in The Immune System - Robert J. BinderDocument186 pagesHeat Shock Proteins in The Immune System - Robert J. BinderHari NugrohoNo ratings yet
- Name Worksheet: Protein and Amino Acid: o o o oDocument2 pagesName Worksheet: Protein and Amino Acid: o o o oShakilaNo ratings yet
- Lista de Precios BCN Insitute AGOSTO 2021Document2 pagesLista de Precios BCN Insitute AGOSTO 2021Estefania SilvaNo ratings yet
- Biochemistry 1st Semester BSN MCQS, Educational Platform ADDocument19 pagesBiochemistry 1st Semester BSN MCQS, Educational Platform ADPrince Masroor Ali Abro100% (1)
- Test Bank For Biochemistry 7th Edition Jeremy M BergDocument6 pagesTest Bank For Biochemistry 7th Edition Jeremy M BergkhuyenhenryzwoowNo ratings yet




























































































Download as pdf or txt
You might also like
- Stoker Chapter19 (Lipids) - 2 PDFDocument85 pagesStoker Chapter19 (Lipids) - 2 PDFAsther Mantua86% (7)
- Experiment 5 LipidsDocument13 pagesExperiment 5 LipidsClemence Marie FuentesNo ratings yet
- 08LipAMO SuarezDocument15 pages08LipAMO SuarezscasuarezNo ratings yet
- Lipid MetabolismDocument32 pagesLipid MetabolismNahid HasanNo ratings yet
- Chapter 08 LipidDocument10 pagesChapter 08 Lipid楊畯凱No ratings yet
- SESE 115 Biochemistry: Don Mariano Marcos Memorial State University La Union PhilippinesDocument13 pagesSESE 115 Biochemistry: Don Mariano Marcos Memorial State University La Union PhilippinesVhenz MapiliNo ratings yet
- Handout 4 - LipidsDocument7 pagesHandout 4 - LipidsPaul CliffordNo ratings yet
- UntitledDocument7 pagesUntitledGodstime sakaNo ratings yet
- Structure of Membrane LipidsDocument41 pagesStructure of Membrane LipidsStanley ChikoveNo ratings yet
- Unit-Iv-Lipids and PorphyrinsDocument29 pagesUnit-Iv-Lipids and PorphyrinsNila ArivoliNo ratings yet
- 05 - Intro UNIT 4Document23 pages05 - Intro UNIT 4Elena OlmedoNo ratings yet
- Biological MacromoleculesDocument2 pagesBiological MacromoleculesBeyoung GamefightNo ratings yet
- Basic Metabolism II Notes Reviewed-1Document80 pagesBasic Metabolism II Notes Reviewed-1felixNo ratings yet
- Chemistry and Digestion of LipidsDocument35 pagesChemistry and Digestion of LipidsBiph BiphNo ratings yet
- BIO 121 Tutorial 14-07-2021Document19 pagesBIO 121 Tutorial 14-07-2021PARVATHY ANIL - IMS20211No ratings yet
- MC2 Biochemistry Lecture Notes For BSN First Semester, 2019-2020 Prepared By: SALINA OSIAL - ALFADDocument5 pagesMC2 Biochemistry Lecture Notes For BSN First Semester, 2019-2020 Prepared By: SALINA OSIAL - ALFADAl-waleed JulkanainNo ratings yet
- CHEM 140 Unit 8 Lec 1 LipidsDocument93 pagesCHEM 140 Unit 8 Lec 1 LipidsMevil Jane MabrasNo ratings yet
- L3 LipidsDocument38 pagesL3 LipidsAman ImranNo ratings yet
- Biochemistry of Lipids PDFDocument51 pagesBiochemistry of Lipids PDFZayan HaiderNo ratings yet
- Different Types of LipidsDocument7 pagesDifferent Types of LipidsNESLIHAN ERSOYAKNo ratings yet
- Biochem Exam 3 Study GuideDocument17 pagesBiochem Exam 3 Study GuideGreg ZhangNo ratings yet
- (Biochem A) 1.2 Lipid Chemistry (Viliran)Document12 pages(Biochem A) 1.2 Lipid Chemistry (Viliran)CaeNo ratings yet
- Lipids BiochemDocument2 pagesLipids BiochemDoc Zay VillafuerteNo ratings yet
- LIPID (Lec) 2019Document7 pagesLIPID (Lec) 2019Joanna Marie TulioNo ratings yet
- 2.1.2 Biological Molecules Part 2Document7 pages2.1.2 Biological Molecules Part 2sjNo ratings yet
- Biological Molecules and EnzymesDocument16 pagesBiological Molecules and EnzymesdebbyhooiNo ratings yet
- 2 LipidsDocument90 pages2 LipidsHasen umerNo ratings yet
- Biological Molecules 2: Carbohydrates and LipidsDocument53 pagesBiological Molecules 2: Carbohydrates and LipidsKyla Cheyenne BustosNo ratings yet
- Bio MoleculesDocument6 pagesBio Moleculesoption 360No ratings yet
- Lipids 2.sabDocument18 pagesLipids 2.sabShujaNo ratings yet
- Fats, Oils, Phospholipids, Waxes, Steroids, Soaps: BL10A HALL 2006 1Document34 pagesFats, Oils, Phospholipids, Waxes, Steroids, Soaps: BL10A HALL 2006 1cbar biologyNo ratings yet
- Smomo Dani and Luvo Bio PresentationDocument3 pagesSmomo Dani and Luvo Bio PresentationGugu MthethwaNo ratings yet
- 2-Biological MoleculesDocument47 pages2-Biological MoleculesaithanjuNo ratings yet
- Lecture 2 Lipids 3Document23 pagesLecture 2 Lipids 3Caamir Dek HaybeNo ratings yet
- Lipids Are A Diverse Group of Hydrophobic MoleculesDocument5 pagesLipids Are A Diverse Group of Hydrophobic MoleculesDanielle GuerraNo ratings yet
- The Chemical Constituents of Cells1Document14 pagesThe Chemical Constituents of Cells1ArnelBautistaNo ratings yet
- Notes On Lipids and Nucleic Acids-Df 19-1Document21 pagesNotes On Lipids and Nucleic Acids-Df 19-1Albert MweembaNo ratings yet
- C46 Lec6 LipidsDocument91 pagesC46 Lec6 LipidsMary Aniderf Cagape Maturan-GumanoyNo ratings yet
- BiomoleculesDocument4 pagesBiomoleculesSchuyler PeytonNo ratings yet
- Module 3 LipidsDocument8 pagesModule 3 LipidsAiza OrdonoNo ratings yet
- Lipids LehningerDocument7 pagesLipids LehningerElla BangalanNo ratings yet
- 10.2 Lipids IntroductionDocument55 pages10.2 Lipids Introductionperssivesimangavwa2002No ratings yet
- Lecture 2 - Introductory BiochemistryDocument15 pagesLecture 2 - Introductory BiochemistryJana-Tae KerrNo ratings yet
- Structure and Classification of Lipids: Lipids Are Naturally Occurring MoleculesDocument19 pagesStructure and Classification of Lipids: Lipids Are Naturally Occurring MoleculesKomic WebtoonNo ratings yet
- Topic-1A (Food and Health) (Autosaved) - 1-20Document20 pagesTopic-1A (Food and Health) (Autosaved) - 1-20lisaNo ratings yet
- BS1001 LipidsDocument32 pagesBS1001 LipidsLiloyi LupiyaNo ratings yet
- BIOMOLECULES EditedDocument4 pagesBIOMOLECULES Editedtgeorgina630No ratings yet
- Pre-Labs 2 Organic Compositions of The CellDocument6 pagesPre-Labs 2 Organic Compositions of The CellTrần Xuân QuỳnhNo ratings yet
- Fatty Acid MetabolismDocument8 pagesFatty Acid MetabolismJessicaOktavianusNo ratings yet
- (Lipid) : Report AboutDocument8 pages(Lipid) : Report AboutMahamad BakrNo ratings yet
- 1.4A Lipid ChemistryDocument12 pages1.4A Lipid ChemistryBea SamonteNo ratings yet
- Chapter 5: Lipids: 5.1. General Considerations About LipidDocument7 pagesChapter 5: Lipids: 5.1. General Considerations About LipidMD. SAJID GHUFRANNo ratings yet
- Biochemistry: Water Carbohydrates Lipids ProteinsDocument16 pagesBiochemistry: Water Carbohydrates Lipids ProteinsryanNo ratings yet
- Digestion and Absorption of LipidsDocument5 pagesDigestion and Absorption of LipidsAngelo Jude Cobacha100% (1)
- Lipids and Nucleic AcidsDocument20 pagesLipids and Nucleic Acidsengr_rimNo ratings yet
- CHO Chemistry: Functions of CarbohydratesDocument3 pagesCHO Chemistry: Functions of CarbohydratesSaadia AnjumNo ratings yet
- Lecture 2 - Introductory BiochemistryDocument17 pagesLecture 2 - Introductory BiochemistryKevonSingh1No ratings yet
- A Level BiologyDocument522 pagesA Level BiologyJAMESNo ratings yet
- 12 - Analysis of LipidsDocument15 pages12 - Analysis of LipidsLIAN KEI HILADONo ratings yet
- Anatomy and Physiology For Students: A College Level Study Guide for Life Science and Allied Health MajorsFrom EverandAnatomy and Physiology For Students: A College Level Study Guide for Life Science and Allied Health MajorsNo ratings yet
- Biochemistry Applied to the Brewing Processes - Fermentation and the Finished BeerFrom EverandBiochemistry Applied to the Brewing Processes - Fermentation and the Finished BeerRating: 5 out of 5 stars5/5 (1)
- C (Dr/Ãfrfi" P¿Z U) HZFT I) LGJL "8L45F86: Lgit Ol Fy (GL H ZL TFZLBMDocument5 pagesC (Dr/Ãfrfi" P¿Z U) HZFT I) LGJL "8L45F86: Lgit Ol Fy (GL H ZL TFZLBMMukeshsinh GadhaviNo ratings yet
- PN 41 2020Document7 pagesPN 41 2020Mukeshsinh GadhaviNo ratings yet
- MA Scheme ExamDocument6 pagesMA Scheme ExamMukeshsinh GadhaviNo ratings yet
- 54 July 1974 Vol. 2 No. 3: Biochemical EducationDocument2 pages54 July 1974 Vol. 2 No. 3: Biochemical EducationMukeshsinh GadhaviNo ratings yet
- Om Prakash PPT BiologyDocument11 pagesOm Prakash PPT BiologyOm Prakash100% (1)
- Desain Tanpa JudulDocument15 pagesDesain Tanpa JudulAprilia NaomiNo ratings yet
- Fixed Oils and Fats: Group 3Document19 pagesFixed Oils and Fats: Group 3Addis MémñøňNo ratings yet
- Nucleic Acid TransDocument2 pagesNucleic Acid TransJae TyNo ratings yet
- Chapter 1 - Part 1 Introduction Classification of Edible OilDocument60 pagesChapter 1 - Part 1 Introduction Classification of Edible OilFirdaus ZubbirNo ratings yet
- Soal CC StrokeDocument1 pageSoal CC StrokeAngeline BudimanNo ratings yet
- Enzyme WorksheetDocument3 pagesEnzyme Worksheetchinmayee chitturu100% (1)
- Arellano University: Physical ScienceDocument22 pagesArellano University: Physical ScienceMa Angelica MasangcayNo ratings yet
- Preparation and Properties of Cheese FudgeDocument10 pagesPreparation and Properties of Cheese FudgeCM MaryNo ratings yet
- Effects of Multi-Enzyme Supplementation On Nutrient Digestibility in Broiler ChickensDocument6 pagesEffects of Multi-Enzyme Supplementation On Nutrient Digestibility in Broiler ChickensOliver TalipNo ratings yet
- BP303TP MergedDocument10 pagesBP303TP MergedManish ChauhanNo ratings yet
- Pcog Assignment Lipids Olfu PharmacyDocument6 pagesPcog Assignment Lipids Olfu Pharmacyadbiral2226cabNo ratings yet
- Prune Juice Concentrate 70 Brix Nutritional InformationDocument2 pagesPrune Juice Concentrate 70 Brix Nutritional Informationborn2dive 9702No ratings yet
- Daksh MahajanDocument12 pagesDaksh MahajanslippypizzaNo ratings yet
- Nonribosomal Peptide Synthetase Biosynthetic Clusters of ESKAPE PathogensDocument29 pagesNonribosomal Peptide Synthetase Biosynthetic Clusters of ESKAPE PathogensMarina DomencoNo ratings yet
- DOPA BiomoleculesDocument9 pagesDOPA Biomoleculesantaryami barikNo ratings yet
- Nucleic AcidsDocument5 pagesNucleic AcidsKheza SuravillaNo ratings yet
- Matdid 207367Document40 pagesMatdid 207367Deblina JanaNo ratings yet
- General Biology 2Document4 pagesGeneral Biology 2Christianne Joseph MonzonNo ratings yet
- Proteins Exam QSDocument5 pagesProteins Exam QSSourav ArunNo ratings yet
- PSmarkup - Jurnal Karya Tulis Ilmiah CiplukanDocument5 pagesPSmarkup - Jurnal Karya Tulis Ilmiah CiplukanrazakNo ratings yet
- Recommended Dietary Allowances - VitaminasDocument3 pagesRecommended Dietary Allowances - VitaminasDaniel Eduardo Lopez CabraNo ratings yet
- animAL Husb. ss2Document1 pageanimAL Husb. ss2Jesse MosesNo ratings yet
- The Central Dogma of Molecular BiologyDocument45 pagesThe Central Dogma of Molecular BiologyEirla Kneriz Trinidad67% (3)
- Heat Shock Proteins in The Immune System - Robert J. BinderDocument186 pagesHeat Shock Proteins in The Immune System - Robert J. BinderHari NugrohoNo ratings yet
- Name Worksheet: Protein and Amino Acid: o o o oDocument2 pagesName Worksheet: Protein and Amino Acid: o o o oShakilaNo ratings yet
- Lista de Precios BCN Insitute AGOSTO 2021Document2 pagesLista de Precios BCN Insitute AGOSTO 2021Estefania SilvaNo ratings yet
- Biochemistry 1st Semester BSN MCQS, Educational Platform ADDocument19 pagesBiochemistry 1st Semester BSN MCQS, Educational Platform ADPrince Masroor Ali Abro100% (1)
- Test Bank For Biochemistry 7th Edition Jeremy M BergDocument6 pagesTest Bank For Biochemistry 7th Edition Jeremy M BergkhuyenhenryzwoowNo ratings yet