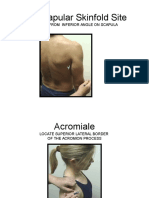
Download as pdf or txt
You might also like
- Human Anatomy 6th Edition Kenneth SaladinDocument11 pagesHuman Anatomy 6th Edition Kenneth SaladinRosennaseNo ratings yet
- ONLINE - Lab 1 - Language of AnatomyDocument9 pagesONLINE - Lab 1 - Language of AnatomyaliNo ratings yet
- Frog Virtual DissectionDocument2 pagesFrog Virtual DissectionLance ZambranoNo ratings yet
- Staghorn Corals of the World: A Revision of the Genus AcroporaFrom EverandStaghorn Corals of the World: A Revision of the Genus AcroporaNo ratings yet
- Lange Radiology of The Chest DiseasesDocument362 pagesLange Radiology of The Chest DiseasesAlexandra Nica100% (1)
- Body Mathching This 1Document6 pagesBody Mathching This 1api-421876727No ratings yet
- Assessment of The Skeletal Pattern and Its Relevance in Treatment PlanningDocument66 pagesAssessment of The Skeletal Pattern and Its Relevance in Treatment PlanningRobert100% (1)
- Lambert 2016Document71 pagesLambert 2016Iqbal FitriantoNo ratings yet
- Phylogeny and Biogeography of Asian Pit VipersDocument32 pagesPhylogeny and Biogeography of Asian Pit VipersHaydee ParasNo ratings yet
- App 20100005Document22 pagesApp 20100005Ghaier KazmiNo ratings yet
- Serpulids (Annelida, Polychaeta) at Cretaceous To Modern Hydrocarbon Seeps Ecological and Evolutionary PatternsDocument7 pagesSerpulids (Annelida, Polychaeta) at Cretaceous To Modern Hydrocarbon Seeps Ecological and Evolutionary PatternsMaria Isabel Aguilar PerezNo ratings yet
- 360 Wake Annurev - Ecolsys 09Document20 pages360 Wake Annurev - Ecolsys 09bex9ggNo ratings yet
- Ascidia World DataBaseDocument5 pagesAscidia World DataBasePatricia UntuNo ratings yet
- Fossil Evidence and The Origin of Bats: Gregg F. Gunnell and Nancy B. SimmonsDocument38 pagesFossil Evidence and The Origin of Bats: Gregg F. Gunnell and Nancy B. SimmonsRenz Junyll ApigoNo ratings yet
- UntitledDocument63 pagesUntitledsanju vNo ratings yet
- A New Rhynchosaur (Reptilia, Archosauromorpha) From The Upper Triassic of Eastern North AmericaDocument12 pagesA New Rhynchosaur (Reptilia, Archosauromorpha) From The Upper Triassic of Eastern North AmericaPatriciaNo ratings yet
- The Lower Jurassic Ornithischian Dinosaur Cranial Anatomy, Functional Morphology, Taxonomy, and RelationshipsDocument95 pagesThe Lower Jurassic Ornithischian Dinosaur Cranial Anatomy, Functional Morphology, Taxonomy, and Relationshipsbraydenstoch777No ratings yet
- Ward 2015 AustralopitecinosDocument14 pagesWard 2015 Australopitecinostomascostelafre1410No ratings yet
- Moon Et Al 2019 Feeding in VertebratesDocument48 pagesMoon Et Al 2019 Feeding in VertebratesMíriam DuarteNo ratings yet
- Serpentes FósseisDocument11 pagesSerpentes FósseisudodrossiNo ratings yet
- Advances in Proboscidean Taxonomy & Classification, Anatomy & Physiology, and Ecology & BehaviorDocument16 pagesAdvances in Proboscidean Taxonomy & Classification, Anatomy & Physiology, and Ecology & BehaviorXp Julieth HernandezNo ratings yet
- A Revision of Australian River Prawns, Macrobrachium, Short 2004Document100 pagesA Revision of Australian River Prawns, Macrobrachium, Short 2004Lutsi SaptaNo ratings yet
- GauthierDocument306 pagesGauthierDan IstrateNo ratings yet
- The University of Chicago Press Wenner-Gren Foundation For Anthropological ResearchDocument22 pagesThe University of Chicago Press Wenner-Gren Foundation For Anthropological ResearchJoel Acuña ZavaletaNo ratings yet
- Craniomandibular Anatomy of A Juvenile Specimen of Edmontosaurus Regalis Lambe 1917 Clarifies Issues in Ontogeny and BiogeographyDocument22 pagesCraniomandibular Anatomy of A Juvenile Specimen of Edmontosaurus Regalis Lambe 1917 Clarifies Issues in Ontogeny and BiogeographyCrocodilo VasconcelosNo ratings yet
- Analysis of Dental Homologies and Phylogeny of Paucituberculata (Mammalia: Marsupialia)Document25 pagesAnalysis of Dental Homologies and Phylogeny of Paucituberculata (Mammalia: Marsupialia)mioclaenusNo ratings yet
- Population Structure of The Gomphothere Stegomastodon Waringi (Mammalia: Proboscidea: Gomphotheriidae) From The Pleistocene of BrazilDocument14 pagesPopulation Structure of The Gomphothere Stegomastodon Waringi (Mammalia: Proboscidea: Gomphotheriidae) From The Pleistocene of BrazilKanekoNo ratings yet
- Sperling and Stockey 2018Document18 pagesSperling and Stockey 2018yoo jjNo ratings yet
- 10 1016@j Geobios 2020 07 005Document27 pages10 1016@j Geobios 2020 07 005DirtyBut WholeNo ratings yet
- 4 PDFDocument11 pages4 PDFmarie parfanNo ratings yet
- Experimental Study of Bone Modification by Captive CaracalDocument13 pagesExperimental Study of Bone Modification by Captive CaracalSidra ZahoorNo ratings yet
- Evidence My Presentation Outline RESUELTADocument7 pagesEvidence My Presentation Outline RESUELTAMartin Emilio Romero BaqueroNo ratings yet
- A Redescription of Chaoyangia Beishanensis Aves and A Comprehensive Phylogeny of Mesozoic BirdsDocument19 pagesA Redescription of Chaoyangia Beishanensis Aves and A Comprehensive Phylogeny of Mesozoic BirdsHector LealNo ratings yet
- Into The Deep: A Phylogenetic Approach To The Bivalve Subclass ProtobranchiaDocument17 pagesInto The Deep: A Phylogenetic Approach To The Bivalve Subclass ProtobranchiaVanessa TumangNo ratings yet
- Deinonychus Hunting 2007 PDFDocument36 pagesDeinonychus Hunting 2007 PDFBibbidyNo ratings yet
- Hou 2014Document39 pagesHou 2014carlos.emr1991No ratings yet
- Alberdi Prado 2022 SAm Gomphothere DiversityDocument8 pagesAlberdi Prado 2022 SAm Gomphothere DiversityLaura Sofia Chinchilla MoralesNo ratings yet
- Miller 2017Document61 pagesMiller 2017Constanza VergaraNo ratings yet
- M Rafi Burhani - 1806137412 - GASTROPODDocument11 pagesM Rafi Burhani - 1806137412 - GASTROPODRAFINo ratings yet
- Bologna Pinto 2002 Old World Genera MeloidaeDocument91 pagesBologna Pinto 2002 Old World Genera MeloidaeoscarbukuNo ratings yet
- 10 Bright OmbarDocument54 pages10 Bright OmbarDbaltNo ratings yet
- Ochotonidae Final ReportDocument15 pagesOchotonidae Final Reportapi-638550362No ratings yet
- Robson 2007 AsianmyrmecologyDocument20 pagesRobson 2007 AsianmyrmecologyRika LauraNo ratings yet
- Pough. Amphibian Biology and HusbandryDocument11 pagesPough. Amphibian Biology and HusbandryJTheron58No ratings yet
- The Ecology of Meiolania Platyceps, A Pleistocene Turtle From AustraliaDocument7 pagesThe Ecology of Meiolania Platyceps, A Pleistocene Turtle From AustraliaHuáscar Bustillos CayojaNo ratings yet
- The Evolution of Early Vertebrate Photoreceptors: ReviewDocument16 pagesThe Evolution of Early Vertebrate Photoreceptors: ReviewJ.D. NobleNo ratings yet
- PINHEIRO Et Al 2013 - A New Eocaiman Alligatoridae Crocodylia From The Itabora Basin Paleogene of Rio de Janeiro BrazilDocument12 pagesPINHEIRO Et Al 2013 - A New Eocaiman Alligatoridae Crocodylia From The Itabora Basin Paleogene of Rio de Janeiro Braziljpmm3535No ratings yet
- YoulaDocument16 pagesYoulacatalinaNo ratings yet
- Vitt Et Al. 2003 - (History and The Global Ecology of Squamate Reptiles)Document17 pagesVitt Et Al. 2003 - (History and The Global Ecology of Squamate Reptiles)Raissa SiqueiraNo ratings yet
- A Global Phylogeny of Apple Snails: Gondwanan Origin, Generic Relationships, and The Influence of Outgroup Choice (Caenogastropoda: Ampullariidae)Document16 pagesA Global Phylogeny of Apple Snails: Gondwanan Origin, Generic Relationships, and The Influence of Outgroup Choice (Caenogastropoda: Ampullariidae)SandraJustinoNo ratings yet
- Peerj 2356Document43 pagesPeerj 2356FABIAN JARED MEDINA BARBANo ratings yet
- Diet and Ecology of Extant and Fossil Wild PigsDocument10 pagesDiet and Ecology of Extant and Fossil Wild PigsMaría EscobarNo ratings yet
- Chapter 20 - MolluscaGastropoda CompressedDocument27 pagesChapter 20 - MolluscaGastropoda Compressedaulia rahmahNo ratings yet
- Ecology and Evolution of Phytotelm-Breeding AnuransDocument84 pagesEcology and Evolution of Phytotelm-Breeding AnuransFabricius Domingos100% (1)
- BarnaclesDocument8 pagesBarnaclesPratyush NahakNo ratings yet
- Artikel Filum MolluscaDocument10 pagesArtikel Filum MolluscaListiyani Putri AzzahroNo ratings yet
- Phylogenetic Relationships and Biogeographic Range Evolution in Cat-Eyed Snakes, Boiga (Serpentes: Colubridae)Document16 pagesPhylogenetic Relationships and Biogeographic Range Evolution in Cat-Eyed Snakes, Boiga (Serpentes: Colubridae)DirtyBut WholeNo ratings yet
- Diversity and Biogeography of Testate AmoebaeDocument15 pagesDiversity and Biogeography of Testate AmoebaeJacky ElíasNo ratings yet
- Re-Description and Range Extension of The Afrotropical Mayfl y Cloeon Perkinsi (Ephemeroptera, Baetidae)Document23 pagesRe-Description and Range Extension of The Afrotropical Mayfl y Cloeon Perkinsi (Ephemeroptera, Baetidae)TommasoNo ratings yet
- 1 PBDocument3 pages1 PBYeison Calizaya MeloNo ratings yet
- Bovinae Fosil RecordsDocument11 pagesBovinae Fosil RecordsradulusNo ratings yet
- The Caudal Skeleton of Extant and Fossil Cyprinodontiform Fishes A Comparative MorphologyDocument20 pagesThe Caudal Skeleton of Extant and Fossil Cyprinodontiform Fishes A Comparative MorphologyAquaponics OaxacaNo ratings yet
- Comparative AnatomyDocument38 pagesComparative AnatomyKimm Charmaine RodriguezNo ratings yet
- Origen HerviboriaDocument263 pagesOrigen HerviboriaLAOGOTIK100% (2)
- Australian Dragonflies: A Guide to the Identification, Distributions and Habitats of Australian OdonataFrom EverandAustralian Dragonflies: A Guide to the Identification, Distributions and Habitats of Australian OdonataRating: 3 out of 5 stars3/5 (1)
- Journal Pre-Proofs: Molecular Phylogenetics and EvolutionDocument21 pagesJournal Pre-Proofs: Molecular Phylogenetics and EvolutionDirtyBut WholeNo ratings yet
- 10 1016@j Geobios 2020 07 005Document27 pages10 1016@j Geobios 2020 07 005DirtyBut WholeNo ratings yet
- ATLAS SERPENTES BRAZIL Nogueira Et Al. 2019 PDFDocument275 pagesATLAS SERPENTES BRAZIL Nogueira Et Al. 2019 PDFrgiorgettiNo ratings yet
- Peerj 10073Document25 pagesPeerj 10073DirtyBut WholeNo ratings yet
- Enigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDocument6 pagesEnigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDirtyBut WholeNo ratings yet
- Bringsoe Et Al. (2020) Oligodon Fasciolatus Eating Organs of Duttaphrynus MelanostictusDocument7 pagesBringsoe Et Al. (2020) Oligodon Fasciolatus Eating Organs of Duttaphrynus MelanostictusDirtyBut WholeNo ratings yet
- Study Ancistrodon Leucostoma: The in WDocument9 pagesStudy Ancistrodon Leucostoma: The in WDirtyBut WholeNo ratings yet
- Bur Brink 2011Document10 pagesBur Brink 2011DirtyBut WholeNo ratings yet
- Phylogenetic Relationships and Biogeographic Range Evolution in Cat-Eyed Snakes, Boiga (Serpentes: Colubridae)Document16 pagesPhylogenetic Relationships and Biogeographic Range Evolution in Cat-Eyed Snakes, Boiga (Serpentes: Colubridae)DirtyBut WholeNo ratings yet
- Introduction To Human Anatomy and PhysiologyDocument21 pagesIntroduction To Human Anatomy and PhysiologyAnghel LopezNo ratings yet
- Examination of The Upper Cervical Spine PDFDocument22 pagesExamination of The Upper Cervical Spine PDFiikemNo ratings yet
- LANDMARKSDocument42 pagesLANDMARKSAndrés Ledezma EncinasNo ratings yet
- AF-Raysofthe WorldredDocument3 pagesAF-Raysofthe WorldredHammurabi RamirezNo ratings yet
- Anatomy AND Physiology: Anatomical PositionDocument9 pagesAnatomy AND Physiology: Anatomical PositionRiyalynkate DellomesNo ratings yet
- Anatomy NotesDocument43 pagesAnatomy NotesLNo ratings yet
- The Pterygomaxillary JunctionDocument11 pagesThe Pterygomaxillary JunctionDONGXU LIUNo ratings yet
- 2018 Automated Three-Dimensional Measurement of Glenoid Version and Inclination in Arthritic ShouldersDocument9 pages2018 Automated Three-Dimensional Measurement of Glenoid Version and Inclination in Arthritic Shouldersusman khanNo ratings yet
- The Human BodyDocument17 pagesThe Human BodyRuthie MendozaNo ratings yet
- Cardio Vascular System of Prawn.Document3 pagesCardio Vascular System of Prawn.Roy AronNo ratings yet
- MCI FMGE Previous Year Solved Question Paper 2003 PDFDocument30 pagesMCI FMGE Previous Year Solved Question Paper 2003 PDFrekha meenaNo ratings yet
- Test Bank For Anatomy Physiology and Disease 2nd Edition ColbertDocument31 pagesTest Bank For Anatomy Physiology and Disease 2nd Edition ColbertJoseph Benson100% (41)
- Van KedisiDocument5 pagesVan KedisiSamil SefergilNo ratings yet
- Lower Lumbar Facet Joint Complex AnatomyDocument8 pagesLower Lumbar Facet Joint Complex AnatomyyohanNo ratings yet
- 1996 19Document5 pages1996 19Anonymous kdBDppigENo ratings yet
- Aptian Albian Rudist Bivalves Hippuritida From The Chilean Central Andes Their Palaeoceanographic Significance 2015 Cretaceous ResearchDocument14 pagesAptian Albian Rudist Bivalves Hippuritida From The Chilean Central Andes Their Palaeoceanographic Significance 2015 Cretaceous ResearchBACLIMERNo ratings yet
- Chapter 1: Introduction To Anatomy and Physiology NotesDocument6 pagesChapter 1: Introduction To Anatomy and Physiology Notesapi-305436791No ratings yet
- OcclusionDocument233 pagesOcclusionLore Dan0% (1)
- Condylar SagDocument8 pagesCondylar Sagshreya dasNo ratings yet
- The Frankel Appliance (FR-2) - Model Preparation and Appliance ConstructionDocument18 pagesThe Frankel Appliance (FR-2) - Model Preparation and Appliance ConstructionMohammed Hussain100% (1)
- PSLD Operation ProcedureDocument2 pagesPSLD Operation ProcedureSonny WijanarkoNo ratings yet
- Anatomy & Physiology I Laboratory Version B: Review Sheet Exercise 1 (The Language of Anatomy)Document5 pagesAnatomy & Physiology I Laboratory Version B: Review Sheet Exercise 1 (The Language of Anatomy)sournsweetNo ratings yet
- Kinesiology of Shoulder ComplexDocument7 pagesKinesiology of Shoulder ComplexAlya Putri KhairaniNo ratings yet
- 1 - Anatomical Terminology KEYDocument6 pages1 - Anatomical Terminology KEY愛.NiniNo ratings yet