Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5825)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Medical TattooingDocument66 pagesMedical TattooingTattooTattoo100% (4)
- Theories and Constructs of RaceDocument16 pagesTheories and Constructs of RaceX69hotdog100% (1)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Chromabright Skin Brightener PresentationDocument22 pagesChromabright Skin Brightener PresentationRnD Roi SuryaNo ratings yet
- Reading Wuiz Hazırlık RE3 1-3Document13 pagesReading Wuiz Hazırlık RE3 1-3Kübra Şendoğan100% (1)
- Signature Assignment Paper Anth 2Document8 pagesSignature Assignment Paper Anth 2api-301347675No ratings yet
- A Guide To Skin Material With ZBrush and Single Pass BPR PabloMunozGDocument29 pagesA Guide To Skin Material With ZBrush and Single Pass BPR PabloMunozGJackes David Lemos Lemos100% (3)
- 406 Melanostatine - Brochure - WebDocument4 pages406 Melanostatine - Brochure - Webapi-291771056No ratings yet
- Basf Presentation Radiant SkinDocument39 pagesBasf Presentation Radiant SkinJames WuNo ratings yet
- StarDesignPower CargillBeauty PresentationDocument28 pagesStarDesignPower CargillBeauty PresentationJames WuNo ratings yet
- Minoxidil Mechanisms of Action On Hair Growthj.1365-2133.2004.05785.x PDFDocument9 pagesMinoxidil Mechanisms of Action On Hair Growthj.1365-2133.2004.05785.x PDFJames WuNo ratings yet
- Chronobiology Biological Clocks and Rhythms of The Skin A. Mehling A J.W. Fluhr BDocument8 pagesChronobiology Biological Clocks and Rhythms of The Skin A. Mehling A J.W. Fluhr BJames WuNo ratings yet
- Treatment of Keloids and Hypertrophic Scars With The Triple-Mode ErYAG Laser A Pilot Study J.mla.2010.10.001Document6 pagesTreatment of Keloids and Hypertrophic Scars With The Triple-Mode ErYAG Laser A Pilot Study J.mla.2010.10.001James WuNo ratings yet
- Perspectives: An Open Introduction To Cultural Anthropology: Second EditionDocument28 pagesPerspectives: An Open Introduction To Cultural Anthropology: Second EditionJy Sie Jé100% (1)
- A "Lost White Tribe" That Created A Global Racial CasteDocument330 pagesA "Lost White Tribe" That Created A Global Racial CasteAsar100% (2)
- Physical Assessment Body Part Normal Findings Actual Findings Interpretation and Analysis NormalDocument24 pagesPhysical Assessment Body Part Normal Findings Actual Findings Interpretation and Analysis Normaljose santiagoNo ratings yet
- HHS Public Access: Color Bar Tool For Skin Type Self-Identification: A Cross Sectional StudyDocument4 pagesHHS Public Access: Color Bar Tool For Skin Type Self-Identification: A Cross Sectional StudyRiefka Ananda ZulfaNo ratings yet
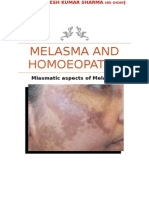
- Melasma and HomoeopathyDocument14 pagesMelasma and HomoeopathyDr. Rajneesh Kumar Sharma MD HomNo ratings yet
- Discussion Guide For Grades 8 & Up The Skin I'm inDocument14 pagesDiscussion Guide For Grades 8 & Up The Skin I'm inMERVE TÜRKNo ratings yet
- Unit 1 Race PaperDocument8 pagesUnit 1 Race Paperapi-266904394No ratings yet
- Increased Blood Flow and Vasculature in Solar LentigoDocument5 pagesIncreased Blood Flow and Vasculature in Solar LentigoOzheanAMNo ratings yet
- Topic 1 Reading Exercises From: Copi, Irving M. Introduction To Logic, 14th Edition. Routledge. Chapter 1Document9 pagesTopic 1 Reading Exercises From: Copi, Irving M. Introduction To Logic, 14th Edition. Routledge. Chapter 1Yash RamawatNo ratings yet
- HCP Problem StatementDocument5 pagesHCP Problem StatementAnonymous MNn0v3No ratings yet
- Comm Studies Ia SpeechDocument3 pagesComm Studies Ia SpeechDaniel McknightNo ratings yet
- Correlating Skin Color With Gingival Pigmentation Patterns in South Indians - A Cross Sectional StudyDocument5 pagesCorrelating Skin Color With Gingival Pigmentation Patterns in South Indians - A Cross Sectional StudyShintia HawariNo ratings yet
- KUKUU-TUNTUM The Ancestral Jurisdiction NhomaDocument81 pagesKUKUU-TUNTUM The Ancestral Jurisdiction NhomabornmagneticNo ratings yet
- Kristen Marrinan-American Beauty StandardsDocument32 pagesKristen Marrinan-American Beauty StandardsMaria RomeroNo ratings yet
- Natriance Brightener - PresentationDocument8 pagesNatriance Brightener - PresentationWootNo ratings yet
- AlbinismDocument10 pagesAlbinismTapash GopeNo ratings yet
- Wagner Et Al., 2002 (Comparing Quantitative Measures)Document6 pagesWagner Et Al., 2002 (Comparing Quantitative Measures)Aurelia SuryaniNo ratings yet
- A Brief History of Filipinos' Obsession With White SkinDocument15 pagesA Brief History of Filipinos' Obsession With White SkinJerico RiveraNo ratings yet
- Mutant Future Section 3 MutationsDocument14 pagesMutant Future Section 3 MutationsMike MitchellNo ratings yet
- Hyperpigmentation of The Skin RevisedDocument4 pagesHyperpigmentation of The Skin RevisedSikandar ZahoorNo ratings yet
- Colonialisms Role in The Success of The Filipino Skin WhiteningDocument12 pagesColonialisms Role in The Success of The Filipino Skin WhiteningJOHN CARLOS GERALDINONo ratings yet
- The GreysDocument37 pagesThe Greyskbk250% (2)
- Polygenic and Monogenic InheritanceDocument5 pagesPolygenic and Monogenic Inheritancekhanmunazah242No ratings yet