Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5823)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Professional Responsibility and AccountabilityDocument5 pagesProfessional Responsibility and Accountabilityjasdeepkaurnagra100% (2)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- 100 Modelos B2 EOI Inglés.Document150 pages100 Modelos B2 EOI Inglés.Hannah Gray100% (2)
- Higher Education Programmes Memorandum: InstructionsDocument24 pagesHigher Education Programmes Memorandum: InstructionsValentia Lindi HoncoNo ratings yet
- Nagle Notes On PharmacologyDocument263 pagesNagle Notes On PharmacologyrelyessNo ratings yet
- Counterfeit Healing of Nuga BestDocument6 pagesCounterfeit Healing of Nuga BestWorks E80% (5)
- SWM Ho Icm 70 10 08 2023Document24 pagesSWM Ho Icm 70 10 08 2023Thukaram McaNo ratings yet
- Pa 112013Document24 pagesPa 112013ThePRESSNewsOnlineNo ratings yet
- Pharmacokinetics in Special PopulationsDocument26 pagesPharmacokinetics in Special PopulationsAnkita banerjeeNo ratings yet
- Awareness of Epilepsy in Pharmacy Students in Different Universities of Karachi, PakistanDocument6 pagesAwareness of Epilepsy in Pharmacy Students in Different Universities of Karachi, PakistanSyeda Eshaal JavaidNo ratings yet
- Breathing ExcersizeDocument35 pagesBreathing Excersizebbmoksh100% (1)
- 1B Approval FormDocument1 page1B Approval FormAgnes G. Arguilles Taladtad100% (1)
- Medical Termination of Pregnancy (MTP) Amendment Act, 2021: Why in NewsDocument4 pagesMedical Termination of Pregnancy (MTP) Amendment Act, 2021: Why in NewsVivek GiriNo ratings yet
- Gracella Irwana - G - Pert 05-06 - Sia - 1Document51 pagesGracella Irwana - G - Pert 05-06 - Sia - 1Gracella IrwanaNo ratings yet
- CHS Barkada Kontra Droga Action PlanDocument3 pagesCHS Barkada Kontra Droga Action PlanMARILYN CONSIGNANo ratings yet
- Acute Abdominal PainDocument11 pagesAcute Abdominal Painone_2_nv100% (1)
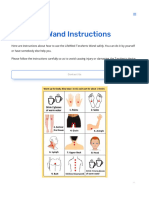
- Terahertz Wand Instructions - LifeWindDocument16 pagesTerahertz Wand Instructions - LifeWindasurya64No ratings yet
- J - A Clinico-Radiographic AnalysisDocument34 pagesJ - A Clinico-Radiographic AnalysisSusovan GiriNo ratings yet
- Tema EnglezaDocument107 pagesTema EnglezaGuvernanta IdealaNo ratings yet
- MSDSDocument11 pagesMSDSkenNo ratings yet
- Biox O2 ProcedimentosDocument9 pagesBiox O2 ProcedimentosClaudia E Luis GonçalvesNo ratings yet
- COVID-19 Vaccine Surveillance Report: Week 1Document54 pagesCOVID-19 Vaccine Surveillance Report: Week 1camaferNo ratings yet
- BIO 230 Syllabus 190 Spring 2024 Angermeier - TaggedDocument9 pagesBIO 230 Syllabus 190 Spring 2024 Angermeier - Taggedkb95387No ratings yet
- Dpco 2013Document17 pagesDpco 2013Padmavathi CNo ratings yet
- 37.2 Agreement Between Employer and MandatoryDocument15 pages37.2 Agreement Between Employer and MandatorykkNo ratings yet
- Total Quality ManagementDocument203 pagesTotal Quality ManagementAnand MosumNo ratings yet
- Asm 2 5047Document36 pagesAsm 2 5047qh11022003No ratings yet
- MDU Indemnity InsuranceDocument2 pagesMDU Indemnity InsuranceMajid Habib KhanNo ratings yet
- Smaw NC IiDocument60 pagesSmaw NC Iisethylijah bernaldezNo ratings yet
- Mental HealthDocument146 pagesMental HealthAnggraeni Beti Dwi LestariNo ratings yet
- CDRMO Assignment byDocument11 pagesCDRMO Assignment byRidaNo ratings yet