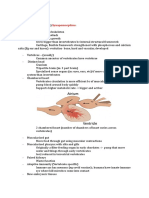
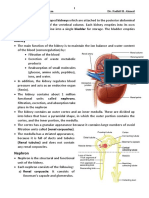
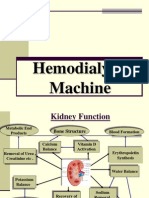
Download as pdf or txt
You might also like
- CSI Guide and Resource Pack: Healthy SkinDocument135 pagesCSI Guide and Resource Pack: Healthy Skinmurse101No ratings yet
- Chapter 5-6 Theories of AgingDocument54 pagesChapter 5-6 Theories of AgingGladz C CadaguitNo ratings yet
- S. Faubel and J. Topf 4 Volume RegulationDocument24 pagesS. Faubel and J. Topf 4 Volume RegulationJoel TopfNo ratings yet
- Lillian BridgesDocument24 pagesLillian BridgesDavidch7100% (5)
- Anatomyoffinfish 180717204712Document36 pagesAnatomyoffinfish 180717204712usriuwi277No ratings yet
- Finand ScaleDocument17 pagesFinand ScalepappuNo ratings yet
- 1 - External Anatomy and TaxonomyDocument8 pages1 - External Anatomy and TaxonomyJuan David Quintero MonsalveNo ratings yet
- Fish ScaleDocument7 pagesFish Scalecrazyleo7450% (2)
- First Semester 2020/2021Document52 pagesFirst Semester 2020/2021low luffecussNo ratings yet
- Fish Anatomy: Ventral Fin Vertical StabilizerDocument16 pagesFish Anatomy: Ventral Fin Vertical Stabilizerlinda_restiNo ratings yet
- Describe The Taxonomical of Identification of Cultivable FishesDocument5 pagesDescribe The Taxonomical of Identification of Cultivable FishesNarasimha MurthyNo ratings yet
- Fins of FishesDocument12 pagesFins of Fishesrano khanNo ratings yet
- Med 8Document2 pagesMed 8Lef RJNo ratings yet
- ) Fusiform: A) Torpedo-Shaped B) Allows Minimal Drag While Swimming C) Best Shape For A Pelagic CruiseDocument63 pages) Fusiform: A) Torpedo-Shaped B) Allows Minimal Drag While Swimming C) Best Shape For A Pelagic CruiseUit CatNo ratings yet
- Lab 5 - ChondreicthyesDocument5 pagesLab 5 - ChondreicthyesCherry Mae AdlawonNo ratings yet
- Fishery NoteDocument14 pagesFishery NoteMegersaNo ratings yet
- Lab 5 - ChondreicthyesDocument5 pagesLab 5 - ChondreicthyesCherry Mae AdlawonNo ratings yet
- SquidDocument6 pagesSquidhaneenNo ratings yet
- Fisheries Biology and Management: External Fish AnatomyDocument7 pagesFisheries Biology and Management: External Fish AnatomyEezem Rachid AushoNo ratings yet
- Jawless FishDocument6 pagesJawless FishmuhammadNo ratings yet
- VertebratesDocument8 pagesVertebratesfeya ayefNo ratings yet
- Bony FishesDocument16 pagesBony FishesmarisNo ratings yet
- Balanoglossus, A Type Representing Protochordata Are Explained in UnitDocument4 pagesBalanoglossus, A Type Representing Protochordata Are Explained in UnitKean GBNo ratings yet
- ShortenDocument18 pagesShortenNick YuNo ratings yet
- Accessory Respiratory Organ - FishDocument27 pagesAccessory Respiratory Organ - Fishkaushik mallick100% (1)
- Vertebrates (Kingdom Animalia)Document130 pagesVertebrates (Kingdom Animalia)Nurl AinaNo ratings yet
- UntitledDocument11 pagesUntitledtaimoor shahzadNo ratings yet
- OSTEICHTHYESDocument2 pagesOSTEICHTHYESMiranda NandaNo ratings yet
- OsteichthyesDocument103 pagesOsteichthyesZaidan Triandi RachmanNo ratings yet
- Fish Classification, StructureDocument40 pagesFish Classification, StructureBio SciencesNo ratings yet
- Growth in FishesDocument353 pagesGrowth in FishesLizviel BragaNo ratings yet
- Exercise 6 Musculo SkeletalDocument9 pagesExercise 6 Musculo SkeletalJacob NoraNo ratings yet
- 4 Sistem-IntegumenDocument49 pages4 Sistem-IntegumenMarius Agustinus WellikenNo ratings yet
- MollusksDocument40 pagesMollusksAlyssa LingamenNo ratings yet
- 4 Sistem-IntegumenDocument49 pages4 Sistem-Integumenchippyz likeNo ratings yet
- Lab 4 - MolluscaDocument18 pagesLab 4 - MolluscaArifameira KinantiNo ratings yet
- Natomy An YDocument2 pagesNatomy An YShawa DabaNo ratings yet
- Text 1 (A & B)Document2 pagesText 1 (A & B)ifx leeNo ratings yet
- FisheriesDocument9 pagesFisheriesSayab DawarNo ratings yet
- Vertebrate Systems SummaryDocument29 pagesVertebrate Systems SummaryHazel Grace Bellen100% (1)
- Structure and Function of Skin of FishDocument11 pagesStructure and Function of Skin of FishBijendra PradhanNo ratings yet
- MolluscsDocument2 pagesMolluscsIvy Villanueva DivinoNo ratings yet
- Fish Skeleto1Document5 pagesFish Skeleto1Ej RafaelNo ratings yet
- Anatomy UmyuDocument8 pagesAnatomy UmyuAminu HassanNo ratings yet
- Description: Elasmobranchii (Document2 pagesDescription: Elasmobranchii (Maui ShihtzuNo ratings yet
- Anatomy of FishDocument11 pagesAnatomy of FishrafumiNo ratings yet
- IFish BiologyDocument42 pagesIFish BiologyJosue Sandigan Biolon SecorinNo ratings yet
- Topic-11 - Mollusca 10Document32 pagesTopic-11 - Mollusca 10Inaya ImranNo ratings yet
- Class OsteichthyesDocument13 pagesClass Osteichthyesedivisha100% (1)
- FishesDocument40 pagesFishesShruti TiwariNo ratings yet
- Bio 342Document76 pagesBio 342Steph VeeNo ratings yet
- ChondrichthyesDocument18 pagesChondrichthyesmey_maya51No ratings yet
- Session 2 Scales of FishesDocument15 pagesSession 2 Scales of FishesAsif KamalNo ratings yet
- Laporan Anatomi Ikan Nilem Dan Ikan LeleDocument9 pagesLaporan Anatomi Ikan Nilem Dan Ikan LeleDaisy KavinskyNo ratings yet
- External Fish AnatomyDocument4 pagesExternal Fish AnatomyKissha Jane100% (1)
- OsteichthyesDocument49 pagesOsteichthyesmuhammadismailNo ratings yet
- Fish Disease Manual 2010 PDFDocument72 pagesFish Disease Manual 2010 PDFvasiliki_eaak100% (2)
- The Phylum Mollusca: A. AmphineuraDocument16 pagesThe Phylum Mollusca: A. AmphineuraCindy RahmahNo ratings yet
- For Fish As Eaten by Humans, See - For Other Uses, See .: From Wikipedia, The Free EncyclopediaDocument24 pagesFor Fish As Eaten by Humans, See - For Other Uses, See .: From Wikipedia, The Free EncyclopediaWira SentanuNo ratings yet
- Mollusc ADocument14 pagesMollusc ADhaffer Al-Mezhany100% (1)
- AEB 302 Lecture NoteDocument92 pagesAEB 302 Lecture Notemisakisuki7No ratings yet
- SciaenidsDocument5 pagesSciaenidsrituNo ratings yet
- Freshwater Aquariums in Your LifeFrom EverandFreshwater Aquariums in Your LifeAmanda PisaniNo ratings yet
- Anim 1014behaviour and Assessment and Management of Welfare in PracticeDocument83 pagesAnim 1014behaviour and Assessment and Management of Welfare in Practicemurse101No ratings yet
- Introduction To Animal WelfareDocument40 pagesIntroduction To Animal Welfaremurse101No ratings yet
- Wound Dressing Guide: Healthy SkinDocument52 pagesWound Dressing Guide: Healthy Skinmurse101No ratings yet
- Positive Behavior Support and Applied Behavior AnalysisDocument24 pagesPositive Behavior Support and Applied Behavior Analysismurse101No ratings yet
- D BirdsDocument45 pagesD Birdsmurse101No ratings yet
- Amphibians: Introduction and EvolutionDocument5 pagesAmphibians: Introduction and Evolutionmurse101No ratings yet
- ESD 1100 Topic 3 Online Lecture 2018Document46 pagesESD 1100 Topic 3 Online Lecture 2018murse101No ratings yet
- Pharmacology of Drugs Acting On Renal SystemDocument20 pagesPharmacology of Drugs Acting On Renal SystemBestha Chakri100% (2)
- Hemodialysis in Stressor Patients - 1Document28 pagesHemodialysis in Stressor Patients - 1Jean Cuenta100% (1)
- Diabetic NephropathyDocument5 pagesDiabetic NephropathyZiedTriki100% (1)
- Kathmandu UniversityDocument65 pagesKathmandu UniversitySurkhali BipanaNo ratings yet
- Chapter - 2: Basic Cell PhysiologyDocument72 pagesChapter - 2: Basic Cell Physiologyyash ingawaleNo ratings yet
- ESPEN Guideline On Clinical Nutrition in Hospitalized Patients With Acute or Chronic Kidney DiseaseDocument25 pagesESPEN Guideline On Clinical Nutrition in Hospitalized Patients With Acute or Chronic Kidney DiseaseOctavianus KevinNo ratings yet
- Duke - AbdominalDocument75 pagesDuke - AbdominalMihir KatlariwalaNo ratings yet
- Hepatorenal Syndrome - UpToDateDocument26 pagesHepatorenal Syndrome - UpToDateAssault AmphibiansNo ratings yet
- Urinary SystemDocument3 pagesUrinary SystemFadhil Hussam AhmedNo ratings yet
- 4800 Week 7 Renal StudentDocument94 pages4800 Week 7 Renal StudentLeAnna100% (1)
- Hepatorrenal PrimerDocument17 pagesHepatorrenal PrimerDianaScandaOlveraCabreraNo ratings yet
- Nefropati DMDocument7 pagesNefropati DMAwaluddin Zheva IdrisNo ratings yet
- How To Dissect RatDocument30 pagesHow To Dissect RatHend MaarofNo ratings yet
- Hemodialysis Machine12Document45 pagesHemodialysis Machine12Divya SoundarajanNo ratings yet
- Clinical Biochemistry Past PaperDocument7 pagesClinical Biochemistry Past PaperAnonymous WkIfo0P100% (1)
- Department of Clinical Biochemistry: Creatinine, SerumDocument3 pagesDepartment of Clinical Biochemistry: Creatinine, SerumrishikundanNo ratings yet
- Crossed and Non-Fused Renal Ectopia of Inferior Type: G. J. Jorwekar, K. N. Dandekar, R. K. Shinde, P. K. BaviskarDocument3 pagesCrossed and Non-Fused Renal Ectopia of Inferior Type: G. J. Jorwekar, K. N. Dandekar, R. K. Shinde, P. K. BaviskarnaseebulNo ratings yet
- MRCP Recall May 19Document9 pagesMRCP Recall May 19Ophthalmology PLUSNo ratings yet
- Renal Embryology: Jason Ryan, MD, MPHDocument731 pagesRenal Embryology: Jason Ryan, MD, MPHFateh BoulounisNo ratings yet
- Insert Kit Kimia KlinikDocument1 pageInsert Kit Kimia Klinikdian saleh100% (1)
- Drug Studies (DMII) (Humulin, NACh03, Losartan, Vessel Due F)Document5 pagesDrug Studies (DMII) (Humulin, NACh03, Losartan, Vessel Due F)Alvin OrizaNo ratings yet
- Safety Assessment and Attenuation of Cisplatin Induced Nephrotoxicity by Tuberous Roots of Boerhaavia DiffusaDocument12 pagesSafety Assessment and Attenuation of Cisplatin Induced Nephrotoxicity by Tuberous Roots of Boerhaavia DiffusaRigotti BrNo ratings yet
- Study of Ocular Manifestations in Patient With Chronic Kidney Disease in Tertiary Care Rural HospitalDocument10 pagesStudy of Ocular Manifestations in Patient With Chronic Kidney Disease in Tertiary Care Rural HospitalIJAR JOURNALNo ratings yet
- Divine Intervention Step 2CK Podcasts Notes - Read Only File - Docx (Dragged) 6 (Dragged) 2Document1 pageDivine Intervention Step 2CK Podcasts Notes - Read Only File - Docx (Dragged) 6 (Dragged) 2winston1234No ratings yet
- Nephrotic Syndrome and Hypertension Nahla I. A L. Gabban Essam Ahmed Abdullah Haider Nadhim Abd FicmsDocument6 pagesNephrotic Syndrome and Hypertension Nahla I. A L. Gabban Essam Ahmed Abdullah Haider Nadhim Abd FicmsMarcelita DuwiriNo ratings yet
- 2020 Крок Англ 1-деньDocument4 pages2020 Крок Англ 1-деньPrince NathanNo ratings yet
- Differential Diagnosis UrolithiasisDocument10 pagesDifferential Diagnosis UrolithiasisAldila Rc X ManiaNo ratings yet