Download as pdf or txt
You might also like
- Basant Agrawal C M Agrawal Engineering DrawingDocument778 pagesBasant Agrawal C M Agrawal Engineering Drawingjay singhvi100% (9)
- Basant Agrawal C M Agrawal Engineering DrawingDocument778 pagesBasant Agrawal C M Agrawal Engineering Drawingjay singhvi100% (9)
- Basant Agrawal C M Agrawal Engineering DrawingDocument778 pagesBasant Agrawal C M Agrawal Engineering Drawingjay singhvi100% (9)
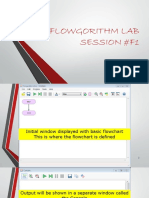
- Flowgorithm PDFDocument56 pagesFlowgorithm PDFKothapalli Likhith Sri SaiNo ratings yet
- Alfred A Barrios - Hypnotherapy A Reappraisal PDFDocument8 pagesAlfred A Barrios - Hypnotherapy A Reappraisal PDFsistemaNo ratings yet
- Challenges of OBDocument10 pagesChallenges of OBGeetanjali Soni 1937629No ratings yet
- Happy Birthday in 100+ LanguagesDocument5 pagesHappy Birthday in 100+ LanguagesRustam Ali0% (1)
- Physiological Mechanisms of Thermoregulation in ReptilesDocument9 pagesPhysiological Mechanisms of Thermoregulation in Reptilesabraao072000No ratings yet
- Physiological Mechanisms of Thermoregulation in ReptilesDocument10 pagesPhysiological Mechanisms of Thermoregulation in Reptilesabraao072000No ratings yet
- Shivering Thermogenesis in Humans Origin, Contribution and MetabolicDocument10 pagesShivering Thermogenesis in Humans Origin, Contribution and MetabolicGio VandaNo ratings yet
- Regulation of Body Temperature by The Nervous System PDFDocument18 pagesRegulation of Body Temperature by The Nervous System PDFEstrella BenitoNo ratings yet
- Animals 14 00513Document21 pagesAnimals 14 00513KhairatulhasanahNo ratings yet
- Trashed-0000318Document5 pagesTrashed-0000318Manish GhimireNo ratings yet
- Central Mechanisms For Thermoregulation in A Hot EnvironmentDocument9 pagesCentral Mechanisms For Thermoregulation in A Hot EnvironmentYonita RahmiNo ratings yet
- Heldmaier Et Al 1989 Photoperiod and Thermoregulation in Vertebrates Body Temperature Rhythms and ThermogenicDocument15 pagesHeldmaier Et Al 1989 Photoperiod and Thermoregulation in Vertebrates Body Temperature Rhythms and Thermogenicsaryasingh253No ratings yet
- 31 Supplement - 5 S157Document5 pages31 Supplement - 5 S157lutfianaNo ratings yet
- On Thermal Sensation and EntropyDocument4 pagesOn Thermal Sensation and Entropyfernando_mart859243No ratings yet
- Fever, Temperature, and The Immune Response: DepartmentDocument12 pagesFever, Temperature, and The Immune Response: DepartmentClara OhNo ratings yet
- Perioperative Shivering ReviewDocument18 pagesPerioperative Shivering Reviewwanoe02No ratings yet
- Being Cool: How Body Temperature Influences Ageing and LongevityDocument15 pagesBeing Cool: How Body Temperature Influences Ageing and LongevityNomalingeNo ratings yet
- Chapter 3 - Intanelina Binti JaluddinDocument4 pagesChapter 3 - Intanelina Binti Jaluddinp2qdfgm7y6No ratings yet
- Neural Substrate of Cold-Seeking Behavior in Endotoxin ShockDocument11 pagesNeural Substrate of Cold-Seeking Behavior in Endotoxin ShockGurumoorthy KrishnamoorthyNo ratings yet
- Update Abiotic StressDocument12 pagesUpdate Abiotic StressYuánArgaNo ratings yet
- Coding Efficiency of Fly Motion Processing Is Set by Firing Rate, Not Firing PrecisionDocument11 pagesCoding Efficiency of Fly Motion Processing Is Set by Firing Rate, Not Firing PrecisionspavieriNo ratings yet
- Molecular and Cellular Endocrinology-Thermoregulation in FishDocument8 pagesMolecular and Cellular Endocrinology-Thermoregulation in FishHAROLD AQUILINO RUIZ MOZOMBITENo ratings yet
- Congeneric Variability in Lifespan Extension and Onset of Senescence Suggest Active Regulation of Aging in Response To Low TemperatureDocument8 pagesCongeneric Variability in Lifespan Extension and Onset of Senescence Suggest Active Regulation of Aging in Response To Low Temperatureender000No ratings yet
- Hipotálamo - Control de La Temperatura - Morrison 2011Document47 pagesHipotálamo - Control de La Temperatura - Morrison 2011fede AssirioNo ratings yet
- Physiological Mechanisms of Thermoregulation in ReptilesDocument9 pagesPhysiological Mechanisms of Thermoregulation in ReptilesSalvador ReyesNo ratings yet
- From Skin to Sensory System. Lumpkin2007Document8 pagesFrom Skin to Sensory System. Lumpkin2007Khang VoNo ratings yet
- Contribution of Thermal and Nonthermal Factors To The Regulation of BodyDocument8 pagesContribution of Thermal and Nonthermal Factors To The Regulation of BodyMike WazowskiNo ratings yet
- Homeostasis and Feedback Mechanism PDFDocument3 pagesHomeostasis and Feedback Mechanism PDFjer montillaNo ratings yet
- Thermoregulation in Vertebrates: Acclimation, Acclimatization and AdaptationDocument8 pagesThermoregulation in Vertebrates: Acclimation, Acclimatization and Adaptationtrunghieu85No ratings yet
- Ajpcell 00040 2004Document11 pagesAjpcell 00040 2004Anibal SalcedoNo ratings yet
- Beyond The Classic Thermoneutral ZoneDocument9 pagesBeyond The Classic Thermoneutral ZonegeoNo ratings yet
- Big 3Document13 pagesBig 3Fitria Ayu LestariNo ratings yet
- TEMPERATURE REGULATION Berne and LevyDocument3 pagesTEMPERATURE REGULATION Berne and LevyMonesa Christy VillanuevaNo ratings yet
- Animals 11 01733 With CoverDocument28 pagesAnimals 11 01733 With CoverNatalia AguasNo ratings yet
- Homeostatic Disturbance of Thermoregulatory Functions in Rats With Chronic FatigueDocument6 pagesHomeostatic Disturbance of Thermoregulatory Functions in Rats With Chronic FatigueG ElegansNo ratings yet
- estrescronicoDepreZORRILLA ZIEHERDocument10 pagesestrescronicoDepreZORRILLA ZIEHERmarielaNo ratings yet
- Variation in The Heat Shock Response and Its Implication For Predicting The Effect of Global Climate Change On Species' Biogeographical Distribution Ranges and Metabolic CostsDocument9 pagesVariation in The Heat Shock Response and Its Implication For Predicting The Effect of Global Climate Change On Species' Biogeographical Distribution Ranges and Metabolic CostspachriqueNo ratings yet
- Acta Physiologica - 2016 - Mrowka - ThermoregulationDocument3 pagesActa Physiologica - 2016 - Mrowka - ThermoregulationSam Skofic (atomichoplite)No ratings yet
- Evaluating Thermoregulation in ReptilesDocument10 pagesEvaluating Thermoregulation in ReptilesSantiago SuarezNo ratings yet
- Lizard ThermoregulationDocument8 pagesLizard Thermoregulationjuan2marquez2No ratings yet
- The Thermoregulatory Theory of Yawning: What We Know From Over 5 Years of ResearchDocument13 pagesThe Thermoregulatory Theory of Yawning: What We Know From Over 5 Years of ResearchAndré AmorimNo ratings yet
- Thermoregulation in BatsDocument45 pagesThermoregulation in BatsBaraa AkeelNo ratings yet
- Suhu Tubuh (Termoregulasi)Document12 pagesSuhu Tubuh (Termoregulasi)Annida LegiNo ratings yet
- Suggestion With AnswerDocument4 pagesSuggestion With AnswerBeautynot2touchNo ratings yet
- Chapperon Et Al 2016Document9 pagesChapperon Et Al 2016Fernando LimaNo ratings yet
- Cavic Chi Oli 2011Document12 pagesCavic Chi Oli 2011Etna VeraguasNo ratings yet
- The Molecular and Cellular Basis of Cold Sensation: David D. MckemyDocument10 pagesThe Molecular and Cellular Basis of Cold Sensation: David D. MckemyAlyanah AloveraNo ratings yet
- Human Thermoregulation and Measurement of Body Temperature in Exercise and Clinical SettingsDocument7 pagesHuman Thermoregulation and Measurement of Body Temperature in Exercise and Clinical SettingsGriffin MulofwaNo ratings yet
- Evidence For Increased Thermogenesis in Female C57Document4 pagesEvidence For Increased Thermogenesis in Female C57Erickson OngNo ratings yet
- 1 s2.0 S2214180419301655 MainDocument7 pages1 s2.0 S2214180419301655 MainMukti WidodoNo ratings yet
- A Newsimplified ThermoregulatorybioheatmodelforevaluatingthermalDocument9 pagesA Newsimplified Thermoregulatorybioheatmodelforevaluatingthermalthi.rar3078No ratings yet
- Tissue 1Document2 pagesTissue 165rangsiman.nanNo ratings yet
- Blouin Demers2005Document7 pagesBlouin Demers2005Rafael Loaiza RamírezNo ratings yet
- Ikegami+et+al Endocrinology Vol156 p647-659Document13 pagesIkegami+et+al Endocrinology Vol156 p647-659sunNo ratings yet
- Microbial ThermosensorsDocument16 pagesMicrobial ThermosensorsClaudia MaturanaNo ratings yet
- s12648-024-03245-wDocument7 pagess12648-024-03245-wHamed BouraouiNo ratings yet
- Thermal Homeostasis in The Newborn Puppy: Behavioral and Physiological ResponsesDocument12 pagesThermal Homeostasis in The Newborn Puppy: Behavioral and Physiological ResponsesHandayaniNo ratings yet
- Parabrachial Neuron Types Categorically Encode Thermoregulation Variables During Heat Defense - 2020Document17 pagesParabrachial Neuron Types Categorically Encode Thermoregulation Variables During Heat Defense - 2020熙槃No ratings yet
- Manuscript SubmitDocument23 pagesManuscript Submitapi-295124915No ratings yet
- Chapter 28-29 PDFDocument45 pagesChapter 28-29 PDFMikayla100% (1)
- Rea T Transfer in Biomedical Engineering and Biotechnology: Proceedings of TheDocument13 pagesRea T Transfer in Biomedical Engineering and Biotechnology: Proceedings of TheHussain Amir AliNo ratings yet
- Environmental_heterogeneity_shapes_physiological_tDocument15 pagesEnvironmental_heterogeneity_shapes_physiological_tkaren hernandezNo ratings yet
- Lecture 11 Temperature and Thermodynamics Part 1 Concepts NotesDocument18 pagesLecture 11 Temperature and Thermodynamics Part 1 Concepts NotesYuuKohNo ratings yet
- 4 - Elementary - Signals Slides - UDocument17 pages4 - Elementary - Signals Slides - UKothapalli Likhith Sri SaiNo ratings yet
- DD Lab ManualDocument12 pagesDD Lab ManualKothapalli Likhith Sri SaiNo ratings yet
- Subspaces of Vector Spaces Math 130 Linear AlgebraDocument2 pagesSubspaces of Vector Spaces Math 130 Linear AlgebraKothapalli Likhith Sri SaiNo ratings yet
- Practice Problems - Metal Drude Model and SemiconductorDocument20 pagesPractice Problems - Metal Drude Model and SemiconductorKothapalli Likhith Sri SaiNo ratings yet
- Ion-Implantation (Very Clear Explanation)Document47 pagesIon-Implantation (Very Clear Explanation)Kothapalli Likhith Sri SaiNo ratings yet
- A Study On Impact of Green Marketing Tools' On PDFDocument41 pagesA Study On Impact of Green Marketing Tools' On PDFSiva VidhyaNo ratings yet
- Rosa Interview Acta SociologicaDocument21 pagesRosa Interview Acta SociologicaDanielNo ratings yet
- Prist University: Question BankDocument4 pagesPrist University: Question BankGanesan NkNo ratings yet
- Thar's Manual of Good Conduct Pages 10 and 11Document1 pageThar's Manual of Good Conduct Pages 10 and 11Grff PritchNo ratings yet
- List of Dragons in Literature - WikipediaDocument10 pagesList of Dragons in Literature - Wikipediajorge solieseNo ratings yet
- Ancestor VenerationDocument43 pagesAncestor VenerationHazel100% (4)
- Lab 1Document3 pagesLab 1milek mathewNo ratings yet
- Portfolio: Learning Activity SheetsDocument23 pagesPortfolio: Learning Activity SheetsMhay Anne Perez100% (1)
- Delhi Domestic Working Womens Forum Vs Union of Ins950517COM455916Document6 pagesDelhi Domestic Working Womens Forum Vs Union of Ins950517COM455916Bleach Body BoiNo ratings yet
- Ejercicios BachilleratoDocument11 pagesEjercicios BachilleratoFilibelen Gonzalez50% (2)
- Heirs of Soledad Alido vs. Flora Campano, or Her Representatives and The Register of Deeds, Province of Iloilo (G.R. No. 226065. July 29, 2019)Document12 pagesHeirs of Soledad Alido vs. Flora Campano, or Her Representatives and The Register of Deeds, Province of Iloilo (G.R. No. 226065. July 29, 2019)HarleneNo ratings yet
- Nebojsa Simin Paper Free Energy of The Oscillating Pendulum-Lever SystemDocument7 pagesNebojsa Simin Paper Free Energy of The Oscillating Pendulum-Lever SystemSith AnanthanNo ratings yet
- Presentación Proyecto Creativo Moderno AzulDocument8 pagesPresentación Proyecto Creativo Moderno AzulValentina HuependoNo ratings yet
- G.R. No. 151895 BOC Vs SerranoDocument4 pagesG.R. No. 151895 BOC Vs SerranoJessie AncogNo ratings yet
- Bahasa Inggris Tentang LingkunganDocument2 pagesBahasa Inggris Tentang LingkunganbukanbasabasiNo ratings yet
- Dissolution of Muslim Marriages Act, 1939Document3 pagesDissolution of Muslim Marriages Act, 1939Lata SinghNo ratings yet
- Fce Cambridge First Certificate Writing Part One Essays Advice and Useful PhrasesDocument5 pagesFce Cambridge First Certificate Writing Part One Essays Advice and Useful PhrasesrNo ratings yet
- Hypotensive Anesthesia in Orthognathic Surgery-A ReviewDocument3 pagesHypotensive Anesthesia in Orthognathic Surgery-A ReviewIOSRjournalNo ratings yet
- 2017 Contamination of Coliform Bacteria in Water and Fishery Resources in Manila Bay Aquaculture FarmsDocument30 pages2017 Contamination of Coliform Bacteria in Water and Fishery Resources in Manila Bay Aquaculture FarmsIsabelNo ratings yet
- ReferencesDocument2 pagesReferencesFakhira ShehzadiNo ratings yet
- MS Access MCQ BankDocument24 pagesMS Access MCQ BankBasavaraju K RNo ratings yet
- 21st Century Literature LESSON LOGDocument2 pages21st Century Literature LESSON LOGEvaNo ratings yet
- Introduction To PsychologyDocument23 pagesIntroduction To PsychologyLa MuérteNo ratings yet
- Nursing Practice Threesoko Na Ito Cgena... Kainis 1Document16 pagesNursing Practice Threesoko Na Ito Cgena... Kainis 1JayCesarNo ratings yet
- Alternative Courses of Action Advantages DisadvantagesDocument3 pagesAlternative Courses of Action Advantages DisadvantagesMaryrose SumulongNo ratings yet
- Cirrhosis and Its Complications Evidence Based TreatmentDocument20 pagesCirrhosis and Its Complications Evidence Based TreatmentSucii Sekar NingrumNo ratings yet
- RPP Spoof WritingDocument9 pagesRPP Spoof WritingPadrin HarlandNo ratings yet