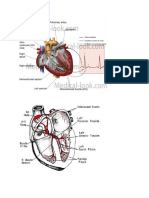
Download as pdf or txt
You might also like
- MRCS A Paper 1 SBAs Applied Basic Sci 2eDocument17 pagesMRCS A Paper 1 SBAs Applied Basic Sci 2eaeages100% (1)
- Anatomy Final Exam Notes PDFDocument45 pagesAnatomy Final Exam Notes PDFLydia LalalandNo ratings yet
- Left Atrial Volume Predicts Cardiovascular EventsDocument6 pagesLeft Atrial Volume Predicts Cardiovascular EventsWirawan PrabowoNo ratings yet
- Physical Exercise Reduces Transplant Arteriosclerosis in A Mouse Aorta Transplantation ModelDocument8 pagesPhysical Exercise Reduces Transplant Arteriosclerosis in A Mouse Aorta Transplantation ModelNicoleta Popa-FoteaNo ratings yet
- Application of Ultrasound in Pulseless Electrical Activity (PEA) Cardiac ArrestDocument3 pagesApplication of Ultrasound in Pulseless Electrical Activity (PEA) Cardiac ArrestKelvin AKNo ratings yet
- 朱家权氯沙坦猪模型Document10 pages朱家权氯沙坦猪模型shentujin1997No ratings yet
- Impaired Left Ventricular Apical Rotation Is Associated With Disease Activity of Psoriatic ArthritisDocument8 pagesImpaired Left Ventricular Apical Rotation Is Associated With Disease Activity of Psoriatic ArthritisEmanuel NavarreteNo ratings yet
- PE and CHFDocument9 pagesPE and CHFKu Li ChengNo ratings yet
- 2016-Clin Nucl Med-Hu-4176787Document3 pages2016-Clin Nucl Med-Hu-4176787LH HuNo ratings yet
- Ehx493 P5438Document1 pageEhx493 P5438Anonymous NeRC5JYiSNo ratings yet
- Anemia in Heart Failure-A Concise ReviewDocument5 pagesAnemia in Heart Failure-A Concise Reviewvikram.vNo ratings yet
- Atrial FibrillationDocument252 pagesAtrial Fibrillationyurikhan100% (1)
- Trepopnea in CHFDocument4 pagesTrepopnea in CHFprobowurNo ratings yet
- Electrocardiographic Artifact MisleadiDocument5 pagesElectrocardiographic Artifact MisleadiMSNo ratings yet
- EcodpplerDocument30 pagesEcodpplerdavidmontoyaNo ratings yet
- Quadricuspid Aortic Valve and A Ventricular Septal Defect in A HorseDocument7 pagesQuadricuspid Aortic Valve and A Ventricular Septal Defect in A HorseJacob HaleNo ratings yet
- CSM 3 2 46 52Document7 pagesCSM 3 2 46 52Santoso 9JimmyNo ratings yet
- Enhancing Detection Accuracy For Clinical Heart Failure Utilizing Pulse Transit Time Variability and Machine LearningDocument9 pagesEnhancing Detection Accuracy For Clinical Heart Failure Utilizing Pulse Transit Time Variability and Machine LearningMuhammad SaimNo ratings yet
- Cardiac ValveDocument9 pagesCardiac Valveikbal rambalinoNo ratings yet
- A Mistake That Has Hurt No One: Sinus MistakusDocument4 pagesA Mistake That Has Hurt No One: Sinus MistakusasclepiuspdfsNo ratings yet
- Genomics and Cardiac Arrhythmias: Cardiovascular Genomic MedicineDocument13 pagesGenomics and Cardiac Arrhythmias: Cardiovascular Genomic MedicineKishore ChinnaNo ratings yet
- Ferric Carboxymaltose in Patients With Heart Failure and Iron DeficiencyDocument13 pagesFerric Carboxymaltose in Patients With Heart Failure and Iron DeficiencyfathaNo ratings yet
- Yesin 2017Document20 pagesYesin 2017Teuku IrfanNo ratings yet
- Moderate To Severe ASDocument7 pagesModerate To Severe ASTheresia Sri RezekiNo ratings yet
- 08 Rao - Evaluation and Management of Wolff-Parkinson-White in Athletes (2014)Document7 pages08 Rao - Evaluation and Management of Wolff-Parkinson-White in Athletes (2014)Yuri medranoNo ratings yet
- Out-Of-hospital Cardiac Arrest - Current Concepts - The Lancet 10marDocument10 pagesOut-Of-hospital Cardiac Arrest - Current Concepts - The Lancet 10marMr. LNo ratings yet
- History WPW ScheinmanDocument6 pagesHistory WPW ScheinmanEdison BonillaNo ratings yet
- Liver Stiffness As Measured by Transient Elastography - 2021 - American Heart JDocument6 pagesLiver Stiffness As Measured by Transient Elastography - 2021 - American Heart JGarret BarriNo ratings yet
- 1 s2.0 S0735109704007946 MainDocument9 pages1 s2.0 S0735109704007946 Mainbagus joyoNo ratings yet
- Through The Decades (Beta) - Blocker Use and Outcomes in Acute Coronary Syndromes.Document8 pagesThrough The Decades (Beta) - Blocker Use and Outcomes in Acute Coronary Syndromes.Tony Miguel Saba SabaNo ratings yet
- VenosoDocument6 pagesVenosocoopervascba.diradmNo ratings yet
- Does This Dyspneic Patient in The Emergency Department Have Congestive Heart Failure?Document13 pagesDoes This Dyspneic Patient in The Emergency Department Have Congestive Heart Failure?Jorge MéndezNo ratings yet
- Doppler en TEPDocument8 pagesDoppler en TEPanytavalenoNo ratings yet
- Stem Cell Therapy An Emerging Treatment Approach For Atrial FibrillationDocument2 pagesStem Cell Therapy An Emerging Treatment Approach For Atrial FibrillationPaul HartingNo ratings yet
- A Cardiac Contouring Atlas For RadiotherapyDocument7 pagesA Cardiac Contouring Atlas For RadiotherapyChendil VNo ratings yet
- Spontaneous Coronary Artery Dissection With Clinical Presentation of Acute Myocardial InfarctionDocument3 pagesSpontaneous Coronary Artery Dissection With Clinical Presentation of Acute Myocardial InfarctionSabrina JonesNo ratings yet
- Cambios en El EKG Predictores de Edema Pulmonar Neurogénico en Hemorragia SubaracnoideaDocument4 pagesCambios en El EKG Predictores de Edema Pulmonar Neurogénico en Hemorragia SubaracnoideaCristina Duran GarcíaNo ratings yet
- Impact of Moderate Aortic Stenosis in Patients With Heart Failure With Reduced Ejection FractionDocument10 pagesImpact of Moderate Aortic Stenosis in Patients With Heart Failure With Reduced Ejection FractionMiguel PugaNo ratings yet
- Etm 14 4 3563 PDFDocument6 pagesEtm 14 4 3563 PDFUlfahNo ratings yet
- Scoring System Identification of High-Risk Patent Foramen Ovale With CSDocument6 pagesScoring System Identification of High-Risk Patent Foramen Ovale With CSStefanie KarinaNo ratings yet
- Dyspnea: How To Differentiate Between Acute Heart Failure Syndrome and Other DiseasesDocument2 pagesDyspnea: How To Differentiate Between Acute Heart Failure Syndrome and Other DiseasesAyaNo ratings yet
- KCJ 42 568Document3 pagesKCJ 42 568vam buddhaNo ratings yet
- Intervesnsi Stroke 1Document9 pagesIntervesnsi Stroke 1Zilbran BerontaxNo ratings yet
- Mckenna 1991Document10 pagesMckenna 1991IsmaelNo ratings yet
- Loukas 2013Document11 pagesLoukas 2013Gabriel Jose CarvajalNo ratings yet
- Hollehorst PlaquesDocument3 pagesHollehorst PlaquesErick Roa GandicaNo ratings yet
- Lin 2009Document7 pagesLin 2009Nur Syamsiah MNo ratings yet
- Xrayand ElectrocardiogramDocument104 pagesXrayand ElectrocardiogramdrelvNo ratings yet
- 1 s2.0 S0894731721000882 MainDocument12 pages1 s2.0 S0894731721000882 MainSuryati HusinNo ratings yet
- Different Clinical Phenotypes of Embolic Stroke of Undetermined Source: A Subgroup Analysis of 86 PatientsDocument9 pagesDifferent Clinical Phenotypes of Embolic Stroke of Undetermined Source: A Subgroup Analysis of 86 PatientsSuryati HusinNo ratings yet
- Di Napoli Et Al 2020 Arterial Spin Labeling Mri in Carotid Stenosis Arterial Transit Artifacts May Predict SymptomsDocument9 pagesDi Napoli Et Al 2020 Arterial Spin Labeling Mri in Carotid Stenosis Arterial Transit Artifacts May Predict SymptomsAdenane BoussoufNo ratings yet
- HF PublishedDocument21 pagesHF Publishedapi-627835898No ratings yet
- Lee 2006Document7 pagesLee 2006Jose Manuel Fuentes Del PozoNo ratings yet
- Article Heart-2001-Lindström-31-8Document9 pagesArticle Heart-2001-Lindström-31-8dgina8800No ratings yet
- Pulmonary Hypertension Is Prevalent in Catheter and Arterio-Venous Access HemodialysisDocument6 pagesPulmonary Hypertension Is Prevalent in Catheter and Arterio-Venous Access HemodialysisMichelle FernandoNo ratings yet
- Untitled PDFDocument2 pagesUntitled PDFmeta ikaNo ratings yet
- Ecor 06 00176Document3 pagesEcor 06 00176saNo ratings yet
- Circulation 1975 Burggraf 146 56Document12 pagesCirculation 1975 Burggraf 146 56Zikri Putra Lan LubisNo ratings yet
- Noc110069 346 351Document6 pagesNoc110069 346 351Carlos AlvaradoNo ratings yet
- Bitter Eur J Heart Fail 2009 11 602Document7 pagesBitter Eur J Heart Fail 2009 11 602Javier Alonso AmaroNo ratings yet
- Clinical Handbook of Cardiac ElectrophysiologyFrom EverandClinical Handbook of Cardiac ElectrophysiologyBenedict M. GloverNo ratings yet
- Cardiovascular System Anatomy and Physiology - Study Guide For NursesDocument34 pagesCardiovascular System Anatomy and Physiology - Study Guide For NursesMarie ParksNo ratings yet
- Possible Essay Questions For The Physiology Final ExamDocument3 pagesPossible Essay Questions For The Physiology Final ExamSalman Majid100% (1)
- RRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRDocument2 pagesRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRRSaginesh MkNo ratings yet
- Electrocardiography II StudentDocument20 pagesElectrocardiography II StudentArmando Valdez ZamoranoNo ratings yet
- The Heart, Part 1 - Under Pressure: Crash Course A&P # 25Document5 pagesThe Heart, Part 1 - Under Pressure: Crash Course A&P # 25Jordan TorresNo ratings yet
- Worksheet Ap06Document7 pagesWorksheet Ap06liawijayaNo ratings yet
- Sistem Konduksi Jantung (Pembekalan) - DR PUT&DrHARDocument32 pagesSistem Konduksi Jantung (Pembekalan) - DR PUT&DrHARNur FarinahNo ratings yet
- Conduction System of The HeartDocument9 pagesConduction System of The Hearthirap50% (2)
- PHS 205 Cardiovascular SystemDocument33 pagesPHS 205 Cardiovascular Systemdivineraymond34No ratings yet
- Physiology of Heart in DetailDocument101 pagesPhysiology of Heart in Detailakanksha sharmaNo ratings yet
- Laporan Kasus 6 Cvs - Kelompok J - 2013Document47 pagesLaporan Kasus 6 Cvs - Kelompok J - 2013Nadiya Afifah AgusNo ratings yet
- Cardiac DysrhythmiasDocument54 pagesCardiac DysrhythmiasClark LlameraNo ratings yet
- Chap. 19 - The Heart (Cardiology)Document96 pagesChap. 19 - The Heart (Cardiology)VARUN SINGHNo ratings yet
- Muthia-Ahmad Basic Cardiac ElectrophysiologyDocument53 pagesMuthia-Ahmad Basic Cardiac ElectrophysiologyFikriYTNo ratings yet
- Mcq-Heart Conducting SystemDocument42 pagesMcq-Heart Conducting SystemDan BrightNo ratings yet
- Martin Biel Et Al - Hyperpolarization-Activated Cation Channels: From Genes To FunctionDocument40 pagesMartin Biel Et Al - Hyperpolarization-Activated Cation Channels: From Genes To FunctionFedrmNo ratings yet
- Effects of Drugs On The Frogs HeartDocument16 pagesEffects of Drugs On The Frogs HeartGreetingwedausezzt50% (2)
- Biology Form 5 Notes Chapter 1Document39 pagesBiology Form 5 Notes Chapter 1Mayghen SelvanayagamNo ratings yet
- Cardiac Action PotentialDocument9 pagesCardiac Action PotentialswennecaNo ratings yet
- Ekg Ecg Interpretation by DR Jason Lowe PDF Bb3 DR NotesDocument79 pagesEkg Ecg Interpretation by DR Jason Lowe PDF Bb3 DR NotesGuru RewriNo ratings yet
- ECG Reading & InterpretationDocument52 pagesECG Reading & InterpretationRoy Francis B. CaspeNo ratings yet
- Practice Worksheets - Cardiac PhysiologyDocument5 pagesPractice Worksheets - Cardiac PhysiologyrickNo ratings yet
- Hye Doun Jan Feb March 08 PSDocument12 pagesHye Doun Jan Feb March 08 PSanrcinfoNo ratings yet
- (PHA) 3.06 Antiarrhythmic Drugs - Dr.Document18 pages(PHA) 3.06 Antiarrhythmic Drugs - Dr.Mickaela Jaden KimNo ratings yet
- Pharm Fall Cardiovascular Pharmacology Study Guide-106Document47 pagesPharm Fall Cardiovascular Pharmacology Study Guide-106sean liyanageNo ratings yet
- Action PotentialDocument24 pagesAction PotentialJohn MarcelNo ratings yet
- The Music of Life-Sourcebook PDFDocument98 pagesThe Music of Life-Sourcebook PDFAmal-krishnadas D'RiveroNo ratings yet
- 07 Human Physiology The Muscular SystemDocument68 pages07 Human Physiology The Muscular SystemsuNo ratings yet