Download as pdf or txt
You might also like
- IGCSE Biology Chapter 1Document42 pagesIGCSE Biology Chapter 1Eric Chew100% (2)
- Oscar Marquez-Martinez - CH 9 QuestionsDocument12 pagesOscar Marquez-Martinez - CH 9 Questionsapi-478637931No ratings yet
- Species Concept, Speciation Evolutionary Thought Before DarwinDocument26 pagesSpecies Concept, Speciation Evolutionary Thought Before DarwinmashedtomatoezedNo ratings yet
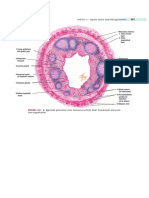
- Histology of AppendixDocument1 pageHistology of Appendixalyson_l100% (1)
- Natural SelectionDocument81 pagesNatural Selectionpatricia piliNo ratings yet
- S Announcement 28055Document115 pagesS Announcement 28055patricia piliNo ratings yet
- Dawkins, Richard - Joint Nesting in A Digger Wasp As An Evolutionarily Stable... (1979)Document44 pagesDawkins, Richard - Joint Nesting in A Digger Wasp As An Evolutionarily Stable... (1979)Jessi GordonNo ratings yet
- P401-01 Microevolutionary Forces - ChapterDocument26 pagesP401-01 Microevolutionary Forces - ChapterBekkem Lavanya ReddyNo ratings yet
- 5.4 EvolutionDocument9 pages5.4 EvolutionIsla HazeltonNo ratings yet
- mODULE 3Document33 pagesmODULE 3Michael Clinton DionedaNo ratings yet
- The Species Concept-1Document5 pagesThe Species Concept-1O me khanNo ratings yet
- Mod 6 Evolution and ClassificationDocument7 pagesMod 6 Evolution and ClassificationrasingtanyaroseNo ratings yet
- Coevolution. Ed. by D.J. Futuyma & M. SlatkinDocument4 pagesCoevolution. Ed. by D.J. Futuyma & M. SlatkinoonthaiNo ratings yet
- Ayala - Teleological Explanations in Evolutionary BiologyDocument16 pagesAyala - Teleological Explanations in Evolutionary BiologyRodrigo CarvalhoNo ratings yet
- BiologyIIforNonMajorsII 02Document69 pagesBiologyIIforNonMajorsII 02GERONA GABRYLE MARCNo ratings yet
- Chapter 4 EvolutionDocument8 pagesChapter 4 EvolutionYuri MiyaNo ratings yet
- Evolution Study GuideDocument7 pagesEvolution Study Guidegmanb5100% (3)
- Teleology BiologyDocument16 pagesTeleology BiologySanil VisalamanaskanNo ratings yet
- Biodiversity 11Document22 pagesBiodiversity 11Ma Naida EndrinalNo ratings yet
- Herre Et Al. 2008Document23 pagesHerre Et Al. 2008Mauricio CostaNo ratings yet
- The Evolution of CooperationDocument27 pagesThe Evolution of Cooperationatilla arıcıoğluNo ratings yet
- Nowak Etal - Evolution of Eusociality - Nature 2010Document49 pagesNowak Etal - Evolution of Eusociality - Nature 2010versoluceNo ratings yet
- Artculo Encyclopedia Evolutionary BiologyDocument9 pagesArtculo Encyclopedia Evolutionary BiologyDaianaNo ratings yet
- Resource Collection RevisedDocument8 pagesResource Collection Revisedapi-691490367No ratings yet
- Insect Biodiversity Perspectives From The SystematDocument3 pagesInsect Biodiversity Perspectives From The SystematkapjukeloketNo ratings yet
- Theories of EvolutionDocument5 pagesTheories of EvolutionMehar FathimaNo ratings yet
- Chapter 5: Interactions in The Ecosystem: 5.1 Habitats and Niches 5.2 Evolution and Adaptation 5.3 PopulationsDocument49 pagesChapter 5: Interactions in The Ecosystem: 5.1 Habitats and Niches 5.2 Evolution and Adaptation 5.3 PopulationsJan Marlon GoritNo ratings yet
- Mechanisms of Population Change: Chapter ConceptsDocument40 pagesMechanisms of Population Change: Chapter ConceptsFrances Ijeoma ObiakorNo ratings yet
- Evolutionary Mechanisms 1) Artificial SelectionDocument5 pagesEvolutionary Mechanisms 1) Artificial SelectionpokesurferNo ratings yet
- Natural Selection and The Origin of Species: Chapter 5 Glossary Worksheet (From Textbook (P87) /PPT)Document4 pagesNatural Selection and The Origin of Species: Chapter 5 Glossary Worksheet (From Textbook (P87) /PPT)Maddah HussainNo ratings yet
- Multilevel Selection: Theoretical Foundations, Historical Examples, and Empirical EvidenceFrom EverandMultilevel Selection: Theoretical Foundations, Historical Examples, and Empirical EvidenceNo ratings yet
- Sympatric Speciation - WikipediaDocument4 pagesSympatric Speciation - Wikipediaskline3No ratings yet
- Analyzing Predator-Prey Models Using Systems of Ordinary Linear D PDFDocument28 pagesAnalyzing Predator-Prey Models Using Systems of Ordinary Linear D PDFAnonymous Zd2zqhDYzQNo ratings yet
- Topic 4 - Biodiversity and Natural Resources PDFDocument11 pagesTopic 4 - Biodiversity and Natural Resources PDFLamia IslamNo ratings yet
- Natural SelectionDocument11 pagesNatural SelectionYuh moddaNo ratings yet
- 18.2 Formation of New Species: Species and The Ability To ReproduceDocument11 pages18.2 Formation of New Species: Species and The Ability To Reproduceklavier10244379No ratings yet
- Claar Et Al. 2021Document20 pagesClaar Et Al. 2021steeven.j.alejandroNo ratings yet
- Charles Darwin and Natural Selection Slide 2 Now That We Have Explored Many of The Processes Involved in Producing and Sustaining Living OrganismsDocument33 pagesCharles Darwin and Natural Selection Slide 2 Now That We Have Explored Many of The Processes Involved in Producing and Sustaining Living OrganismsurdanetacalNo ratings yet
- Biological AltruismDocument16 pagesBiological AltruismmehdiukNo ratings yet
- Evolutionary Ecology across Three Trophic Levels: Goldenrods, Gallmakers, and Natural Enemies (MPB-29)From EverandEvolutionary Ecology across Three Trophic Levels: Goldenrods, Gallmakers, and Natural Enemies (MPB-29)Rating: 4 out of 5 stars4/5 (1)
- ESS Assessment Prep PDFDocument7 pagesESS Assessment Prep PDFareenaNo ratings yet
- Assignment 2Document4 pagesAssignment 2Cameron TragesserNo ratings yet
- Ereshefsky Species Pluralism and AntirealismDocument19 pagesEreshefsky Species Pluralism and AntirealismCallum Woods-StrideNo ratings yet
- Exam 1-EcologyDocument7 pagesExam 1-EcologymaiagottholdNo ratings yet
- Unit8L1-4 SCIENCEDocument8 pagesUnit8L1-4 SCIENCEKhrysNo ratings yet
- Hypothesis of Sympatric Speciation 2Document4 pagesHypothesis of Sympatric Speciation 2Saeedullah SeoNo ratings yet
- TELEOLOGICAL EXPLANATIONS IN EVOLUTIONARY BIOLOGY. by FRANCISCODocument16 pagesTELEOLOGICAL EXPLANATIONS IN EVOLUTIONARY BIOLOGY. by FRANCISCOTrust IssuesNo ratings yet
- Understanding Evolution: Gene Selection vs. Group Selection: Michaela LewisDocument3 pagesUnderstanding Evolution: Gene Selection vs. Group Selection: Michaela Lewisnugroho_why_youNo ratings yet
- Lesson 6 EvolutionDocument61 pagesLesson 6 EvolutionCRIS EDISON VILLANUEVANo ratings yet
- Envi. Sci. Final TermDocument39 pagesEnvi. Sci. Final Termmesadaeterjohn.studentNo ratings yet
- Grade 9 Science Biological Diversity Final Exam Preparation: Study GuideDocument14 pagesGrade 9 Science Biological Diversity Final Exam Preparation: Study GuideJelly BaldonadoNo ratings yet
- Aggregation and Predator Avoidance in Whirligig Beetles (Coleoptera: Gyrinidae)Document12 pagesAggregation and Predator Avoidance in Whirligig Beetles (Coleoptera: Gyrinidae)Guy ZerNo ratings yet
- How Interaction Between Organism and Their Environment Leads To Evolution and AdaptationDocument10 pagesHow Interaction Between Organism and Their Environment Leads To Evolution and AdaptationGimme Your WafflesNo ratings yet
- Teoria Nicho ArticuloDocument7 pagesTeoria Nicho ArticuloANA LUCIA CORTES LOAIZANo ratings yet
- Introduction Class 1Document10 pagesIntroduction Class 1an23ms268No ratings yet
- Altig Johnston1989 PDFDocument30 pagesAltig Johnston1989 PDFITALO ALVARENGA GONCALVESNo ratings yet
- Science 8 - Q4 - Week 4Document29 pagesScience 8 - Q4 - Week 4AlfielAquino100% (1)
- Genetic Variation and Polymorphim in The Inducible Spines of A Marine BryozoanDocument13 pagesGenetic Variation and Polymorphim in The Inducible Spines of A Marine BryozoanCarlos MeirellesNo ratings yet
- Nama: Citra Ariza 1911060269 Dwi Rahma Pelita 1911060286 Kelas: Biologi 4DDocument7 pagesNama: Citra Ariza 1911060269 Dwi Rahma Pelita 1911060286 Kelas: Biologi 4Ddwi rahmaNo ratings yet
- ATQ (Bio Lab 12) E7Document3 pagesATQ (Bio Lab 12) E7fizzypurplesodaNo ratings yet
- Topic 8. Evolution Biological Communities and Species InteractionsDocument15 pagesTopic 8. Evolution Biological Communities and Species Interactions科琳No ratings yet
- Novel Aggregation Pheromone, (1S, 4R) - P-Menth-2-En-1-Ol, of The Ambrosia Beetle, Platypus Quercivorus (Coleoptera: Platypodidae)Document10 pagesNovel Aggregation Pheromone, (1S, 4R) - P-Menth-2-En-1-Ol, of The Ambrosia Beetle, Platypus Quercivorus (Coleoptera: Platypodidae)heimat66No ratings yet
- Budiewi Et Al 2020Document10 pagesBudiewi Et Al 2020heimat66No ratings yet
- Platypus Sulcatus ( Mutatus) To World Poplar: The Threat of The Ambrosia Beetle, ResourcesDocument3 pagesPlatypus Sulcatus ( Mutatus) To World Poplar: The Threat of The Ambrosia Beetle, Resourcesheimat66No ratings yet
- Allmen Et Al 2004Document4 pagesAllmen Et Al 2004heimat66No ratings yet
- Identification of A Putative Aggregation Pheromone From Males Platypus Cylindrus (Coleoptera: Platypodidae)Document3 pagesIdentification of A Putative Aggregation Pheromone From Males Platypus Cylindrus (Coleoptera: Platypodidae)heimat66No ratings yet
- Aikens Et Al 2013 PDFDocument19 pagesAikens Et Al 2013 PDFheimat66No ratings yet
- The Phylogeny of The Homoptera1: Quick Links To Online ContentDocument19 pagesThe Phylogeny of The Homoptera1: Quick Links To Online Contentheimat66No ratings yet
- Classification of Herbicide Mode of Action HRACDocument7 pagesClassification of Herbicide Mode of Action HRACDavid Panéz50% (4)
- B.I Sec 4.Document18 pagesB.I Sec 4.dark SpizroNo ratings yet
- Amylase Enzyme and Temperature LabDocument4 pagesAmylase Enzyme and Temperature LabJames DaurayNo ratings yet
- Sharmaine Ginete - Laboratory Activity 3Document3 pagesSharmaine Ginete - Laboratory Activity 3Sharmaine GineteNo ratings yet
- Sri Chaitanya Grand Test - 03Document24 pagesSri Chaitanya Grand Test - 03Subhabrata Mabhai100% (3)
- Cell TransportDocument22 pagesCell TransportxCutex XwolfiechanXNo ratings yet
- BradfordDocument3 pagesBradfordKL TongsonNo ratings yet
- Vitamin C: A. B. C. D. eDocument3 pagesVitamin C: A. B. C. D. ebegoNo ratings yet
- Darlene Krizian G. Dayuha A8A56: Bio100 - General BotanyDocument2 pagesDarlene Krizian G. Dayuha A8A56: Bio100 - General BotanyDarlene KrizianNo ratings yet
- AP BioDocument12 pagesAP BioThomas Lim100% (2)
- Pengaruh Aplikasi Asam Giberelin (Ga3) Terhadap Hasil Benih Padi HibridaDocument8 pagesPengaruh Aplikasi Asam Giberelin (Ga3) Terhadap Hasil Benih Padi HibridaFernando CanggihNo ratings yet
- Plant Disease Epidemiology: PLP 3205 Horticultural Plant PathologyDocument27 pagesPlant Disease Epidemiology: PLP 3205 Horticultural Plant PathologyAddinaNo ratings yet
- Earth and Life Science Week 3-4 Key AnswerDocument18 pagesEarth and Life Science Week 3-4 Key Answerkrysfer03No ratings yet
- 7 STD Chapter 12 Answer KeyDocument1 page7 STD Chapter 12 Answer KeyDeven BariNo ratings yet
- General Heart Diagram, Handout Heart Diagram, Vessels HandoutDocument4 pagesGeneral Heart Diagram, Handout Heart Diagram, Vessels HandoutJNo ratings yet
- Circulatory System: Transporting Gases, Nutrients, Wastes, and HormonesDocument49 pagesCirculatory System: Transporting Gases, Nutrients, Wastes, and HormonesChristopher EstevezNo ratings yet
- Gastrointestinal Masterclass Session 1Document38 pagesGastrointestinal Masterclass Session 1expandNo ratings yet
- Bio Midterm Last YearDocument8 pagesBio Midterm Last YearYun JiNo ratings yet
- Alternative Culture Media For Bacterial Growth Using Different Formulation of Protein SourcesDocument4 pagesAlternative Culture Media For Bacterial Growth Using Different Formulation of Protein SourcesMasitha Dwi AmiraNo ratings yet
- Proteomics Plant StressDocument167 pagesProteomics Plant StressSurajit BhattacharjeeNo ratings yet
- Pathology SEQ Answers - Adaptive Responses & Cell InjuryDocument7 pagesPathology SEQ Answers - Adaptive Responses & Cell InjurysugandiNo ratings yet
- Disease of Immune System: Sharon Hazel Joyce C. Sebastian, RMTDocument47 pagesDisease of Immune System: Sharon Hazel Joyce C. Sebastian, RMTGilo IlaganNo ratings yet
- AgeLOC Y Span Product Brochure LTO ProcessDocument7 pagesAgeLOC Y Span Product Brochure LTO ProcessKent FanNo ratings yet
- EvolutionDocument21 pagesEvolutionLawrence AnDrew FrondaNo ratings yet
- Nsejs OcapsDocument1 pageNsejs Ocapsprem sinhaNo ratings yet
- Oncology Nursing HandoutsDocument7 pagesOncology Nursing HandoutsShandz de Rosas100% (1)
- Chapter 01 - Anatomy and Electrophysiology of The Heart: Multiple Choice QuestionsDocument17 pagesChapter 01 - Anatomy and Electrophysiology of The Heart: Multiple Choice QuestionsyebegashetNo ratings yet
- Ge6351 PDFDocument202 pagesGe6351 PDFrahulNo ratings yet