
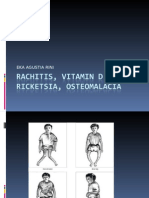
Rickets: Figures 1b and 2
Rickets: Figures 1b and 2
You might also like
- Maternity Case 7: Carla Hernandez (Core) : Guided Reflection QuestionsDocument2 pagesMaternity Case 7: Carla Hernandez (Core) : Guided Reflection QuestionsDai Nguyen100% (1)
- DR Amanullah Lail: Assistant Professor Paediatrics Duhs/Dmc/ChkDocument36 pagesDR Amanullah Lail: Assistant Professor Paediatrics Duhs/Dmc/ChkpaqardNo ratings yet
- Chapter 9Document13 pagesChapter 9Zaky DavidiaNo ratings yet
- Fat Soluble Vitamin RevisedDocument80 pagesFat Soluble Vitamin RevisedsumitNo ratings yet
- 1 RicketsDocument32 pages1 RicketsDarina Ismakaieva100% (1)
- Diseases of Bone MangalaDocument246 pagesDiseases of Bone MangalaharshiniNo ratings yet
- Pediatrics: Vitamin D Deficiency (Rickets) Vitamin D MetabolismDocument3 pagesPediatrics: Vitamin D Deficiency (Rickets) Vitamin D Metabolismapi-3829364No ratings yet
- Lec.5 RicketsDocument7 pagesLec.5 Ricketsبركان معتصم مطشرNo ratings yet
- PEDIATRICSDocument70 pagesPEDIATRICSMathewos MekonnenNo ratings yet
- Fibrous DysplasiaDocument2 pagesFibrous DysplasiaGus SwanjayaNo ratings yet
- Vitamin D MetabolismDocument38 pagesVitamin D MetabolismDanielle Nicole AtienzaNo ratings yet
- CHAPTER 63 and 64 Vitamin CDocument41 pagesCHAPTER 63 and 64 Vitamin CJosselle Sempio CalientaNo ratings yet
- Rickets: Dr. Yasna Kibria MD Resident (Phase A) Department of Radiology and Imaging BsmmuDocument43 pagesRickets: Dr. Yasna Kibria MD Resident (Phase A) Department of Radiology and Imaging BsmmuYasna KibriaNo ratings yet
- Rickets PDFDocument39 pagesRickets PDFLidia Iglesias SiglerNo ratings yet
- Rickets and Hypervitaminosis D: by Dr. NjugunaDocument7 pagesRickets and Hypervitaminosis D: by Dr. NjugunaDavid NjugunaNo ratings yet
- Disorders of Fat Soluble VitaminsDocument22 pagesDisorders of Fat Soluble Vitaminsbpt2No ratings yet
- Metabolic Bone DiseaseDocument36 pagesMetabolic Bone DiseaseArun KumarNo ratings yet
- Watch A Film About Rickets (Causes, Symptoms, Treatment) :: VocabularyDocument6 pagesWatch A Film About Rickets (Causes, Symptoms, Treatment) :: VocabularyKorin MijaNo ratings yet
- Vitamin Deficiency KvsDocument67 pagesVitamin Deficiency KvsVarsini C PNo ratings yet
- Osteochondritis Dissecans in The Dog: Issue 1 Article 12Document7 pagesOsteochondritis Dissecans in The Dog: Issue 1 Article 12rizky holijaNo ratings yet
- Down's Syndrome and Spina BifidaDocument23 pagesDown's Syndrome and Spina BifidaKajal KaliaNo ratings yet
- Intro: Marie & Sainton's Disease, Scheuthaner-Marie-Saniton Syndrome, Mutational DystosisDocument4 pagesIntro: Marie & Sainton's Disease, Scheuthaner-Marie-Saniton Syndrome, Mutational DystosisNoor-E-Khadiza ShamaNo ratings yet
- A Practical Approach To Diagnose and Treat Rickets: Journal of Clinical Medicine of Kazakhstan (E-ISSN 2313-1519)Document7 pagesA Practical Approach To Diagnose and Treat Rickets: Journal of Clinical Medicine of Kazakhstan (E-ISSN 2313-1519)goppalNo ratings yet
- Lumbar Spinal Stenosis: Devin K. Binder, M.D., PH.D, Meic H. Schmidt, M.D., and Philip R. Weinstein, M.DDocument10 pagesLumbar Spinal Stenosis: Devin K. Binder, M.D., PH.D, Meic H. Schmidt, M.D., and Philip R. Weinstein, M.DYerni DaeliNo ratings yet
- 01 Metabolic Diseases CADocument47 pages01 Metabolic Diseases CAMi PatelNo ratings yet
- Fibroosseous LesionsDocument28 pagesFibroosseous LesionsDr'Paramjot KaurNo ratings yet
- Httpsamoodle - Su.edu - Egpluginfile.php202864mod Resourcecontent1Lecture20120 Bone20diseas20 PDFDocument7 pagesHttpsamoodle - Su.edu - Egpluginfile.php202864mod Resourcecontent1Lecture20120 Bone20diseas20 PDFtthtn6c8pbNo ratings yet
- 11 Fibro Osseous LesionsDocument22 pages11 Fibro Osseous Lesionsفراس الموسويNo ratings yet
- Spina Bifida General Dental PracticeDocument5 pagesSpina Bifida General Dental PracticerambuanggieNo ratings yet
- 1 - Vitamins A, B1, B3 & CDocument8 pages1 - Vitamins A, B1, B3 & CIbrahim RamizNo ratings yet
- Fibrous Dysplasia: Kepaniteraan Klinik Senior Bagian Ilmu Bedah Orthopaedic Royal Prima MedanDocument14 pagesFibrous Dysplasia: Kepaniteraan Klinik Senior Bagian Ilmu Bedah Orthopaedic Royal Prima MedanPutraNo ratings yet
- Metabolic & Endocrine DisorderDocument117 pagesMetabolic & Endocrine DisorderMd.AlauddinNo ratings yet
- Chapter 22: Generalized Disease Conditions of BoneDocument6 pagesChapter 22: Generalized Disease Conditions of BonepoddataNo ratings yet
- Osteogenesis ImperfectaDocument55 pagesOsteogenesis ImperfectaNorman RiyadiNo ratings yet
- Nutritional Rickets and Osteomalacia in The Twenty-First Century: Revised Concepts, Public Health, and Prevention StrategiesDocument10 pagesNutritional Rickets and Osteomalacia in The Twenty-First Century: Revised Concepts, Public Health, and Prevention StrategiesSyeda MariamNo ratings yet
- Word HereditereyDocument7 pagesWord Hereditereypxb8dhqgs9No ratings yet
- Fibrous DysplasiaDocument29 pagesFibrous Dysplasiadr_adjie100% (2)
- Bones & Joints Pathology 4thDocument8 pagesBones & Joints Pathology 4thlovelyc95No ratings yet
- Bahan Tutor 5.2Document7 pagesBahan Tutor 5.2star shipNo ratings yet
- Assignment On Bone DiseaseDocument9 pagesAssignment On Bone DiseaseShamaNo ratings yet
- New Microsoft Word DocumentDocument3 pagesNew Microsoft Word DocumentmeedoasadelNo ratings yet
- Assignment On Bone DiseaseDocument9 pagesAssignment On Bone DiseaseNoor-E-Khadiza ShamaNo ratings yet
- Hetal Patel GROUP - 631: RicketsDocument28 pagesHetal Patel GROUP - 631: RicketsRahul MunikrishnaNo ratings yet
- Disorders of Bone and Mineral Other Than OsteoporosisDocument15 pagesDisorders of Bone and Mineral Other Than OsteoporosisOdilia MariaNo ratings yet
- Vitamin D CalciumDocument6 pagesVitamin D CalciumSajibNo ratings yet
- Sindrome Desostose Cleidocraniana-Trat OrtodDocument8 pagesSindrome Desostose Cleidocraniana-Trat Ortodbeh.chaves16No ratings yet
- Biology ProjectDocument8 pagesBiology ProjectSANJAYNo ratings yet
- Oral Pathology Lec - 1Document9 pagesOral Pathology Lec - 1مصطفى محمدNo ratings yet
- Kuliah Rickets PBLDocument43 pagesKuliah Rickets PBLNovasuryatiNo ratings yet
- Metabolic Bone Diseases: Dr. R.C.JindalDocument73 pagesMetabolic Bone Diseases: Dr. R.C.JindalpriyankNo ratings yet
- ىDocument16 pagesىjack jonNo ratings yet
- Metabolic Jaw DiseasesDocument55 pagesMetabolic Jaw DiseasesoladunniNo ratings yet
- RickricketsDocument30 pagesRickricketsmurali_bharadwazNo ratings yet
- MuscolskeletalDocument74 pagesMuscolskeletalsamar yousif mohamedNo ratings yet
- 2 5336943370080096042 PDFDocument21 pages2 5336943370080096042 PDFعبدالرحمن عبدالسلام ذنون المصريNo ratings yet
- Genetic Bone DiseasesDocument41 pagesGenetic Bone DiseasesKifamulusi ErisaNo ratings yet
- Kuliah OiDocument61 pagesKuliah OiNovasuryatiNo ratings yet
- Fibro-Osseous Lesions of The JawsDocument68 pagesFibro-Osseous Lesions of The JawsAmisha SaxenaNo ratings yet
- Leg Bowing 1Document42 pagesLeg Bowing 1Mitulsinh M RavaljiNo ratings yet
- The Dental Pulp: Biology, Pathology, and Regenerative TherapiesFrom EverandThe Dental Pulp: Biology, Pathology, and Regenerative TherapiesNo ratings yet
- Vaginal Discharge in DogsDocument5 pagesVaginal Discharge in DogsDarmawan Prastyo100% (1)
- Fish and Aquaculture: Executive SummaryDocument10 pagesFish and Aquaculture: Executive SummaryTan SoNo ratings yet
- Coevolution of Hosts and ParasitesDocument5 pagesCoevolution of Hosts and ParasitesAshok KoyiNo ratings yet
- Abus LightweightDocument2 pagesAbus LightweightGloria RomicNo ratings yet
- 1000 English Collocations EbookDocument128 pages1000 English Collocations EbookHerdeiro DicaprioNo ratings yet
- Sinif Ingilizce 8. Unite Calisma Kagidi Test Etkinlik Worksheet FITNESS by Murat DukkanciDocument13 pagesSinif Ingilizce 8. Unite Calisma Kagidi Test Etkinlik Worksheet FITNESS by Murat DukkanciElif yurtsevenNo ratings yet
- Oxidation of N-Acetyl Alanine by Chloramine-T in Presence of Hydrochloric Acid: A Kinetic and Mechanistic StudyDocument10 pagesOxidation of N-Acetyl Alanine by Chloramine-T in Presence of Hydrochloric Acid: A Kinetic and Mechanistic StudyCentral Asian StudiesNo ratings yet
- Styro FlexDocument7 pagesStyro Flexprabhat singhNo ratings yet
- A Review On The Enhancement of Figure of MeritDocument23 pagesA Review On The Enhancement of Figure of Meritsasa_22No ratings yet
- IPC Medium-Pressure Internal Gear Pumps: Technical Data SheetDocument20 pagesIPC Medium-Pressure Internal Gear Pumps: Technical Data SheetNutrición SaludableNo ratings yet
- FSO - 21 - Safety Functions UM - D - A5Document444 pagesFSO - 21 - Safety Functions UM - D - A5Sanu YoNo ratings yet
- SBBK PKM MotongkadDocument56 pagesSBBK PKM MotongkadBayu Firmansyah AlitNo ratings yet
- A La Carte MenuDocument1 pageA La Carte Menuhappy frankensteinNo ratings yet
- Winkler, W & Pelosi - 2022 - Understanding Breast CancerDocument76 pagesWinkler, W & Pelosi - 2022 - Understanding Breast CancerJordan KopongNo ratings yet
- Biology Paper Ib: Maple Leaf International SchoolDocument22 pagesBiology Paper Ib: Maple Leaf International SchoolFcgfht ghkhkujk100% (1)
- Surfactant FloodingDocument13 pagesSurfactant FloodingVlassis SarantinosNo ratings yet
- WPS-PQR RepairDocument16 pagesWPS-PQR RepairmohammedetaNo ratings yet
- SITREP NUMBER 62 As of 29 January 2020 1100HDocument67 pagesSITREP NUMBER 62 As of 29 January 2020 1100HEdison ManaloNo ratings yet
- P4 - Rolled TrufflesDocument4 pagesP4 - Rolled TrufflesWendy JacksonNo ratings yet
- Acids, Bases and Salt Preparations 1 QPDocument8 pagesAcids, Bases and Salt Preparations 1 QPAbdullah SheikhNo ratings yet
- From Gathering To Growing Food: Neinuo's LunchDocument10 pagesFrom Gathering To Growing Food: Neinuo's Lunchsoumya KavdiaNo ratings yet
- Individual Health Insurance Policy CISDocument3 pagesIndividual Health Insurance Policy CISGIJONo ratings yet
- PM Handbook (Full PDFDocument88 pagesPM Handbook (Full PDFGary LoNo ratings yet
- FingerprintingDocument36 pagesFingerprintingapi-291003259100% (2)
- Heridity: Table of SpecificationsDocument8 pagesHeridity: Table of SpecificationsJoseph GratilNo ratings yet
- Nanoscience 2 PDFDocument388 pagesNanoscience 2 PDFSergio ArturoNo ratings yet
- Global Inequality and PovertyDocument16 pagesGlobal Inequality and PovertyEsmaeil ZeinyNo ratings yet
- Building Construction Company ProfileDocument62 pagesBuilding Construction Company ProfileFadhlan Dwi Gusti WarmanNo ratings yet
- UD150L-40E Ope M501-E053GDocument164 pagesUD150L-40E Ope M501-E053GMahmoud Mady100% (3)

























































































Download as pdf or txt
You might also like
- Maternity Case 7: Carla Hernandez (Core) : Guided Reflection QuestionsDocument2 pagesMaternity Case 7: Carla Hernandez (Core) : Guided Reflection QuestionsDai Nguyen100% (1)
- DR Amanullah Lail: Assistant Professor Paediatrics Duhs/Dmc/ChkDocument36 pagesDR Amanullah Lail: Assistant Professor Paediatrics Duhs/Dmc/ChkpaqardNo ratings yet
- Chapter 9Document13 pagesChapter 9Zaky DavidiaNo ratings yet
- Fat Soluble Vitamin RevisedDocument80 pagesFat Soluble Vitamin RevisedsumitNo ratings yet
- 1 RicketsDocument32 pages1 RicketsDarina Ismakaieva100% (1)
- Diseases of Bone MangalaDocument246 pagesDiseases of Bone MangalaharshiniNo ratings yet
- Pediatrics: Vitamin D Deficiency (Rickets) Vitamin D MetabolismDocument3 pagesPediatrics: Vitamin D Deficiency (Rickets) Vitamin D Metabolismapi-3829364No ratings yet
- Lec.5 RicketsDocument7 pagesLec.5 Ricketsبركان معتصم مطشرNo ratings yet
- PEDIATRICSDocument70 pagesPEDIATRICSMathewos MekonnenNo ratings yet
- Fibrous DysplasiaDocument2 pagesFibrous DysplasiaGus SwanjayaNo ratings yet
- Vitamin D MetabolismDocument38 pagesVitamin D MetabolismDanielle Nicole AtienzaNo ratings yet
- CHAPTER 63 and 64 Vitamin CDocument41 pagesCHAPTER 63 and 64 Vitamin CJosselle Sempio CalientaNo ratings yet
- Rickets: Dr. Yasna Kibria MD Resident (Phase A) Department of Radiology and Imaging BsmmuDocument43 pagesRickets: Dr. Yasna Kibria MD Resident (Phase A) Department of Radiology and Imaging BsmmuYasna KibriaNo ratings yet
- Rickets PDFDocument39 pagesRickets PDFLidia Iglesias SiglerNo ratings yet
- Rickets and Hypervitaminosis D: by Dr. NjugunaDocument7 pagesRickets and Hypervitaminosis D: by Dr. NjugunaDavid NjugunaNo ratings yet
- Disorders of Fat Soluble VitaminsDocument22 pagesDisorders of Fat Soluble Vitaminsbpt2No ratings yet
- Metabolic Bone DiseaseDocument36 pagesMetabolic Bone DiseaseArun KumarNo ratings yet
- Watch A Film About Rickets (Causes, Symptoms, Treatment) :: VocabularyDocument6 pagesWatch A Film About Rickets (Causes, Symptoms, Treatment) :: VocabularyKorin MijaNo ratings yet
- Vitamin Deficiency KvsDocument67 pagesVitamin Deficiency KvsVarsini C PNo ratings yet
- Osteochondritis Dissecans in The Dog: Issue 1 Article 12Document7 pagesOsteochondritis Dissecans in The Dog: Issue 1 Article 12rizky holijaNo ratings yet
- Down's Syndrome and Spina BifidaDocument23 pagesDown's Syndrome and Spina BifidaKajal KaliaNo ratings yet
- Intro: Marie & Sainton's Disease, Scheuthaner-Marie-Saniton Syndrome, Mutational DystosisDocument4 pagesIntro: Marie & Sainton's Disease, Scheuthaner-Marie-Saniton Syndrome, Mutational DystosisNoor-E-Khadiza ShamaNo ratings yet
- A Practical Approach To Diagnose and Treat Rickets: Journal of Clinical Medicine of Kazakhstan (E-ISSN 2313-1519)Document7 pagesA Practical Approach To Diagnose and Treat Rickets: Journal of Clinical Medicine of Kazakhstan (E-ISSN 2313-1519)goppalNo ratings yet
- Lumbar Spinal Stenosis: Devin K. Binder, M.D., PH.D, Meic H. Schmidt, M.D., and Philip R. Weinstein, M.DDocument10 pagesLumbar Spinal Stenosis: Devin K. Binder, M.D., PH.D, Meic H. Schmidt, M.D., and Philip R. Weinstein, M.DYerni DaeliNo ratings yet
- 01 Metabolic Diseases CADocument47 pages01 Metabolic Diseases CAMi PatelNo ratings yet
- Fibroosseous LesionsDocument28 pagesFibroosseous LesionsDr'Paramjot KaurNo ratings yet
- Httpsamoodle - Su.edu - Egpluginfile.php202864mod Resourcecontent1Lecture20120 Bone20diseas20 PDFDocument7 pagesHttpsamoodle - Su.edu - Egpluginfile.php202864mod Resourcecontent1Lecture20120 Bone20diseas20 PDFtthtn6c8pbNo ratings yet
- 11 Fibro Osseous LesionsDocument22 pages11 Fibro Osseous Lesionsفراس الموسويNo ratings yet
- Spina Bifida General Dental PracticeDocument5 pagesSpina Bifida General Dental PracticerambuanggieNo ratings yet
- 1 - Vitamins A, B1, B3 & CDocument8 pages1 - Vitamins A, B1, B3 & CIbrahim RamizNo ratings yet
- Fibrous Dysplasia: Kepaniteraan Klinik Senior Bagian Ilmu Bedah Orthopaedic Royal Prima MedanDocument14 pagesFibrous Dysplasia: Kepaniteraan Klinik Senior Bagian Ilmu Bedah Orthopaedic Royal Prima MedanPutraNo ratings yet
- Metabolic & Endocrine DisorderDocument117 pagesMetabolic & Endocrine DisorderMd.AlauddinNo ratings yet
- Chapter 22: Generalized Disease Conditions of BoneDocument6 pagesChapter 22: Generalized Disease Conditions of BonepoddataNo ratings yet
- Osteogenesis ImperfectaDocument55 pagesOsteogenesis ImperfectaNorman RiyadiNo ratings yet
- Nutritional Rickets and Osteomalacia in The Twenty-First Century: Revised Concepts, Public Health, and Prevention StrategiesDocument10 pagesNutritional Rickets and Osteomalacia in The Twenty-First Century: Revised Concepts, Public Health, and Prevention StrategiesSyeda MariamNo ratings yet
- Word HereditereyDocument7 pagesWord Hereditereypxb8dhqgs9No ratings yet
- Fibrous DysplasiaDocument29 pagesFibrous Dysplasiadr_adjie100% (2)
- Bones & Joints Pathology 4thDocument8 pagesBones & Joints Pathology 4thlovelyc95No ratings yet
- Bahan Tutor 5.2Document7 pagesBahan Tutor 5.2star shipNo ratings yet
- Assignment On Bone DiseaseDocument9 pagesAssignment On Bone DiseaseShamaNo ratings yet
- New Microsoft Word DocumentDocument3 pagesNew Microsoft Word DocumentmeedoasadelNo ratings yet
- Assignment On Bone DiseaseDocument9 pagesAssignment On Bone DiseaseNoor-E-Khadiza ShamaNo ratings yet
- Hetal Patel GROUP - 631: RicketsDocument28 pagesHetal Patel GROUP - 631: RicketsRahul MunikrishnaNo ratings yet
- Disorders of Bone and Mineral Other Than OsteoporosisDocument15 pagesDisorders of Bone and Mineral Other Than OsteoporosisOdilia MariaNo ratings yet
- Vitamin D CalciumDocument6 pagesVitamin D CalciumSajibNo ratings yet
- Sindrome Desostose Cleidocraniana-Trat OrtodDocument8 pagesSindrome Desostose Cleidocraniana-Trat Ortodbeh.chaves16No ratings yet
- Biology ProjectDocument8 pagesBiology ProjectSANJAYNo ratings yet
- Oral Pathology Lec - 1Document9 pagesOral Pathology Lec - 1مصطفى محمدNo ratings yet
- Kuliah Rickets PBLDocument43 pagesKuliah Rickets PBLNovasuryatiNo ratings yet
- Metabolic Bone Diseases: Dr. R.C.JindalDocument73 pagesMetabolic Bone Diseases: Dr. R.C.JindalpriyankNo ratings yet
- ىDocument16 pagesىjack jonNo ratings yet
- Metabolic Jaw DiseasesDocument55 pagesMetabolic Jaw DiseasesoladunniNo ratings yet
- RickricketsDocument30 pagesRickricketsmurali_bharadwazNo ratings yet
- MuscolskeletalDocument74 pagesMuscolskeletalsamar yousif mohamedNo ratings yet
- 2 5336943370080096042 PDFDocument21 pages2 5336943370080096042 PDFعبدالرحمن عبدالسلام ذنون المصريNo ratings yet
- Genetic Bone DiseasesDocument41 pagesGenetic Bone DiseasesKifamulusi ErisaNo ratings yet
- Kuliah OiDocument61 pagesKuliah OiNovasuryatiNo ratings yet
- Fibro-Osseous Lesions of The JawsDocument68 pagesFibro-Osseous Lesions of The JawsAmisha SaxenaNo ratings yet
- Leg Bowing 1Document42 pagesLeg Bowing 1Mitulsinh M RavaljiNo ratings yet
- The Dental Pulp: Biology, Pathology, and Regenerative TherapiesFrom EverandThe Dental Pulp: Biology, Pathology, and Regenerative TherapiesNo ratings yet
- Vaginal Discharge in DogsDocument5 pagesVaginal Discharge in DogsDarmawan Prastyo100% (1)
- Fish and Aquaculture: Executive SummaryDocument10 pagesFish and Aquaculture: Executive SummaryTan SoNo ratings yet
- Coevolution of Hosts and ParasitesDocument5 pagesCoevolution of Hosts and ParasitesAshok KoyiNo ratings yet
- Abus LightweightDocument2 pagesAbus LightweightGloria RomicNo ratings yet
- 1000 English Collocations EbookDocument128 pages1000 English Collocations EbookHerdeiro DicaprioNo ratings yet
- Sinif Ingilizce 8. Unite Calisma Kagidi Test Etkinlik Worksheet FITNESS by Murat DukkanciDocument13 pagesSinif Ingilizce 8. Unite Calisma Kagidi Test Etkinlik Worksheet FITNESS by Murat DukkanciElif yurtsevenNo ratings yet
- Oxidation of N-Acetyl Alanine by Chloramine-T in Presence of Hydrochloric Acid: A Kinetic and Mechanistic StudyDocument10 pagesOxidation of N-Acetyl Alanine by Chloramine-T in Presence of Hydrochloric Acid: A Kinetic and Mechanistic StudyCentral Asian StudiesNo ratings yet
- Styro FlexDocument7 pagesStyro Flexprabhat singhNo ratings yet
- A Review On The Enhancement of Figure of MeritDocument23 pagesA Review On The Enhancement of Figure of Meritsasa_22No ratings yet
- IPC Medium-Pressure Internal Gear Pumps: Technical Data SheetDocument20 pagesIPC Medium-Pressure Internal Gear Pumps: Technical Data SheetNutrición SaludableNo ratings yet
- FSO - 21 - Safety Functions UM - D - A5Document444 pagesFSO - 21 - Safety Functions UM - D - A5Sanu YoNo ratings yet
- SBBK PKM MotongkadDocument56 pagesSBBK PKM MotongkadBayu Firmansyah AlitNo ratings yet
- A La Carte MenuDocument1 pageA La Carte Menuhappy frankensteinNo ratings yet
- Winkler, W & Pelosi - 2022 - Understanding Breast CancerDocument76 pagesWinkler, W & Pelosi - 2022 - Understanding Breast CancerJordan KopongNo ratings yet
- Biology Paper Ib: Maple Leaf International SchoolDocument22 pagesBiology Paper Ib: Maple Leaf International SchoolFcgfht ghkhkujk100% (1)
- Surfactant FloodingDocument13 pagesSurfactant FloodingVlassis SarantinosNo ratings yet
- WPS-PQR RepairDocument16 pagesWPS-PQR RepairmohammedetaNo ratings yet
- SITREP NUMBER 62 As of 29 January 2020 1100HDocument67 pagesSITREP NUMBER 62 As of 29 January 2020 1100HEdison ManaloNo ratings yet
- P4 - Rolled TrufflesDocument4 pagesP4 - Rolled TrufflesWendy JacksonNo ratings yet
- Acids, Bases and Salt Preparations 1 QPDocument8 pagesAcids, Bases and Salt Preparations 1 QPAbdullah SheikhNo ratings yet
- From Gathering To Growing Food: Neinuo's LunchDocument10 pagesFrom Gathering To Growing Food: Neinuo's Lunchsoumya KavdiaNo ratings yet
- Individual Health Insurance Policy CISDocument3 pagesIndividual Health Insurance Policy CISGIJONo ratings yet
- PM Handbook (Full PDFDocument88 pagesPM Handbook (Full PDFGary LoNo ratings yet
- FingerprintingDocument36 pagesFingerprintingapi-291003259100% (2)
- Heridity: Table of SpecificationsDocument8 pagesHeridity: Table of SpecificationsJoseph GratilNo ratings yet
- Nanoscience 2 PDFDocument388 pagesNanoscience 2 PDFSergio ArturoNo ratings yet
- Global Inequality and PovertyDocument16 pagesGlobal Inequality and PovertyEsmaeil ZeinyNo ratings yet
- Building Construction Company ProfileDocument62 pagesBuilding Construction Company ProfileFadhlan Dwi Gusti WarmanNo ratings yet
- UD150L-40E Ope M501-E053GDocument164 pagesUD150L-40E Ope M501-E053GMahmoud Mady100% (3)