Download as pdf or txt
You might also like
- Elements of Ecology 9th Edition Smith Test BankDocument18 pagesElements of Ecology 9th Edition Smith Test Bankjennifercortezmcnpsjtiek100% (32)
- Biology: a QuickStudy Laminated Reference GuideFrom EverandBiology: a QuickStudy Laminated Reference GuideRating: 3 out of 5 stars3/5 (2)
- Essential Biochemistry 3rd Edition - PrattDocument12 pagesEssential Biochemistry 3rd Edition - PrattTaymiah MahmoodNo ratings yet
- Fish Physiology 2001 Vol 19 Tuna Physiology, Ecology, and EvolutionDocument474 pagesFish Physiology 2001 Vol 19 Tuna Physiology, Ecology, and Evolutionlucas17No ratings yet
- General - Biology - II - Q2 Module 5Document20 pagesGeneral - Biology - II - Q2 Module 5Sherry Macalalad Garcia50% (2)
- Cogollodo - Finals Seatwork (Gen Physio)Document2 pagesCogollodo - Finals Seatwork (Gen Physio)Shaira CogollodoNo ratings yet
- Art Sense Sensing The Arts in The Everyday Page 127 132.Document7 pagesArt Sense Sensing The Arts in The Everyday Page 127 132.Shaira CogollodoNo ratings yet
- Art Sense Sensing The Arts in The Everyday 108 113Document6 pagesArt Sense Sensing The Arts in The Everyday 108 113Shaira CogollodoNo ratings yet
- Comparative Animal Physiology: Philip C. WithersDocument7 pagesComparative Animal Physiology: Philip C. WithersWwwanand111No ratings yet
- New DocumentDocument10 pagesNew DocumentIommi Arabella Paras AltreNo ratings yet
- Hipotálamo - Control de La Temperatura - Morrison 2011Document47 pagesHipotálamo - Control de La Temperatura - Morrison 2011fede AssirioNo ratings yet
- Physiological Mechanisms of Thermoregulation in ReptilesDocument10 pagesPhysiological Mechanisms of Thermoregulation in Reptilesabraao072000No ratings yet
- Physiological Mechanisms of Thermoregulation in ReptilesDocument9 pagesPhysiological Mechanisms of Thermoregulation in Reptilesabraao072000No ratings yet
- Tissue 1Document2 pagesTissue 165rangsiman.nanNo ratings yet
- The Origin of Mammalian Endothermy A Paradigm For The Evolution of Complex Biological StructureDocument16 pagesThe Origin of Mammalian Endothermy A Paradigm For The Evolution of Complex Biological StructureWilsonObrigadoNo ratings yet
- Malyshev2016 Article BudDormancyBreakingAffectsRespDocument8 pagesMalyshev2016 Article BudDormancyBreakingAffectsRespjuan antNo ratings yet
- Minireview Heat Shock ProteinsDocument4 pagesMinireview Heat Shock Proteinsnrf2No ratings yet
- Distribuição Da Imunorreatividade Da Proteína de Ligação À TransferrinaDocument12 pagesDistribuição Da Imunorreatividade Da Proteína de Ligação À TransferrinaJose Mauricio Da Silva FerreiraNo ratings yet
- Lesson 1: Introduction To Bioenergetics: Review QuestionsDocument2 pagesLesson 1: Introduction To Bioenergetics: Review QuestionsBea SamonteNo ratings yet
- Evidencia de Un Condicionamiento Conductual en Una AmebaDocument12 pagesEvidencia de Un Condicionamiento Conductual en Una AmebaBellucciNo ratings yet
- 1 - Give An Illustrated Account of The Fine Staracture of A Plant Cell and Describe The Function of The Diffrent Cell OrgnagesDocument19 pages1 - Give An Illustrated Account of The Fine Staracture of A Plant Cell and Describe The Function of The Diffrent Cell OrgnagesMD shah KhanNo ratings yet
- Pys PDFDocument5 pagesPys PDFMD shah KhanNo ratings yet
- Honeybee Colony Thermoregulation - Regulatory Mechanisms and Contribution of Individuals in Dependence On Age, Location and Thermal StressDocument13 pagesHoneybee Colony Thermoregulation - Regulatory Mechanisms and Contribution of Individuals in Dependence On Age, Location and Thermal Stresslosaron437No ratings yet
- Activation of Bacterial Spores. A Review': I G RminaDocument7 pagesActivation of Bacterial Spores. A Review': I G RminaJunegreg CualNo ratings yet
- TMP 2 C4Document8 pagesTMP 2 C4FrontiersNo ratings yet
- Bio 102 End SemDocument177 pagesBio 102 End Sematharvajoshi23No ratings yet
- Prigogine 1972Document7 pagesPrigogine 1972Aleestra100% (1)
- A New Approach To Neural Cell Culture For Long-Term Studies: Steve M. Potter, Thomas B. DemarseDocument8 pagesA New Approach To Neural Cell Culture For Long-Term Studies: Steve M. Potter, Thomas B. DemarsesdfoiuhjwerNo ratings yet
- Comparative Genomic Analysis of The Thermophilic Biomass-Degrading Fungi Myceliophthora Thermophila and Thielavia TerrestrisDocument8 pagesComparative Genomic Analysis of The Thermophilic Biomass-Degrading Fungi Myceliophthora Thermophila and Thielavia TerrestrisDaniel RestrepoNo ratings yet
- Extremophilic Microorganisms: Biochemical Adaptation and Biotechnological Application (Review)Document14 pagesExtremophilic Microorganisms: Biochemical Adaptation and Biotechnological Application (Review)Arely PradoNo ratings yet
- Homeostasis and Feedback Mechanism PDFDocument3 pagesHomeostasis and Feedback Mechanism PDFjer montillaNo ratings yet
- Respiration in Plants: Negros Oriental State UniversityDocument18 pagesRespiration in Plants: Negros Oriental State UniversityGenelie Abuzo CadayonaNo ratings yet
- ThermodynamicsDocument7 pagesThermodynamicsYong YangNo ratings yet
- Specific Learning Objectives - Test 1Document8 pagesSpecific Learning Objectives - Test 1MeenkusNo ratings yet
- Tattersall2016 Seasonal Endothermy ReptilDocument8 pagesTattersall2016 Seasonal Endothermy ReptilAlex RodriguezNo ratings yet
- Lesson1 2Document4 pagesLesson1 2Cheradee AnimNo ratings yet
- 1 s2.0 0166223689901653 MainDocument8 pages1 s2.0 0166223689901653 MainDavid MrosekNo ratings yet
- 30-Second Biochemistry - The 50 Vital Processes in and Around Living Organisms, Each ExplainedDocument162 pages30-Second Biochemistry - The 50 Vital Processes in and Around Living Organisms, Each ExplainedNiaNo ratings yet
- Review Notes2Document31 pagesReview Notes2Diether Añonuevo DavidNo ratings yet
- Bacterial Tyrosinases 1Document12 pagesBacterial Tyrosinases 1Kevin Anthony Oré MaldonadoNo ratings yet
- IGCSE-Biology Paper 1Document91 pagesIGCSE-Biology Paper 1michaela menzelNo ratings yet
- General Biology 2Document4 pagesGeneral Biology 2Charmaine LustriaNo ratings yet
- Essential Chemistry For BiochemistsDocument27 pagesEssential Chemistry For BiochemistsMaria MichaliaNo ratings yet
- Actin 2002Document11 pagesActin 2002Mira45No ratings yet
- 6C Decomposition and Forensics 6C CheckpointDocument6 pages6C Decomposition and Forensics 6C CheckpointsalmaNo ratings yet
- (Setchell, 2006) The Effects of Heat On The Testes of MammalsDocument11 pages(Setchell, 2006) The Effects of Heat On The Testes of MammalsMaiko DantasNo ratings yet
- Neural Substrate of Cold-Seeking Behavior in Endotoxin ShockDocument11 pagesNeural Substrate of Cold-Seeking Behavior in Endotoxin ShockGurumoorthy KrishnamoorthyNo ratings yet
- Biology Notes Paper 1 (Finnished)Document92 pagesBiology Notes Paper 1 (Finnished)michaela menzelNo ratings yet
- Suggestion With AnswerDocument4 pagesSuggestion With AnswerBeautynot2touchNo ratings yet
- Microbial ThermosensorsDocument16 pagesMicrobial ThermosensorsClaudia MaturanaNo ratings yet
- Peroxisomes Glyoxysomes and Glycosomes ReviewDocument14 pagesPeroxisomes Glyoxysomes and Glycosomes ReviewHugo JimenezNo ratings yet
- Mitochondria and Chloroplasts Energy Synthesis of ATP Extracellular ATPDocument4 pagesMitochondria and Chloroplasts Energy Synthesis of ATP Extracellular ATPGiancarlo CasabarNo ratings yet
- Electrorelease of Escherichia Coli NucleoidsDocument6 pagesElectrorelease of Escherichia Coli Nucleoidserhan6936No ratings yet
- MODULE 3 - General ScienceDocument3 pagesMODULE 3 - General ScienceMae GarciaNo ratings yet
- General Biology I ReviewerDocument2 pagesGeneral Biology I ReviewerKathleen Kaye Delos AngelesNo ratings yet
- MITOCHONDRIONDocument6 pagesMITOCHONDRIONCheradee AnimNo ratings yet
- Mammalian Hibernation: Differential Gene Expression and Novel Application of Epigenetic ControlsDocument12 pagesMammalian Hibernation: Differential Gene Expression and Novel Application of Epigenetic ControlsaufaaNo ratings yet
- Physics and Evolution of Thermophilic Adaptation: Igor N. Berezovsky and Eugene I. ShakhnovichDocument6 pagesPhysics and Evolution of Thermophilic Adaptation: Igor N. Berezovsky and Eugene I. ShakhnovichLuthfi NurpratamaNo ratings yet
- Temperature Stress and Responses of Plants: Anna Źróbek-SokolnikDocument22 pagesTemperature Stress and Responses of Plants: Anna Źróbek-SokolnikFatima ZahraNo ratings yet
- My Notes For Blg1502!1!1Document10 pagesMy Notes For Blg1502!1!1Pasipanodya MuzendaNo ratings yet
- Bio 100Document25 pagesBio 100Mariel De Leon GinesNo ratings yet
- Fever - Pathogenesis, Pathophysiology, and PurposeDocument10 pagesFever - Pathogenesis, Pathophysiology, and PurposeTiago RiosNo ratings yet
- Botany ReviewerDocument6 pagesBotany ReviewerAgatha RoseNo ratings yet
- ch7 11 PDFDocument105 pagesch7 11 PDFa portNo ratings yet
- Cell Mol Finals ReviewerDocument11 pagesCell Mol Finals Reviewerswifthomeworks100No ratings yet
- Gen Bio Q1Document11 pagesGen Bio Q1jessy silva (Lynsy)No ratings yet
- Seatwork 2Document3 pagesSeatwork 2Karyl GregorioNo ratings yet
- Here Is Where Your Presentation BeginsDocument39 pagesHere Is Where Your Presentation BeginsShaira CogollodoNo ratings yet
- Finals Gen Physio Lab ExamDocument3 pagesFinals Gen Physio Lab ExamShaira CogollodoNo ratings yet
- Lesson 3 Designing and Managing ServicesDocument26 pagesLesson 3 Designing and Managing ServicesShaira Cogollodo100% (1)
- Improving Service Quality and ProductivityDocument24 pagesImproving Service Quality and ProductivityShaira CogollodoNo ratings yet
- Get To Know Your Customers: Slide ©2004 by Christopher Lovelock and Jochen Wirtz Services Marketing 5/EDocument25 pagesGet To Know Your Customers: Slide ©2004 by Christopher Lovelock and Jochen Wirtz Services Marketing 5/EShaira CogollodoNo ratings yet
- Multiple Choice Questions (MCQ) Topic Quiz Photosynthesis: Instructions and Answers For TeachersDocument29 pagesMultiple Choice Questions (MCQ) Topic Quiz Photosynthesis: Instructions and Answers For TeachersShaira CogollodoNo ratings yet
- White Blue Green and Yellow Photo Collage Modern New Hire Resources Company PresentationDocument9 pagesWhite Blue Green and Yellow Photo Collage Modern New Hire Resources Company PresentationShaira CogollodoNo ratings yet
- Article 1Document12 pagesArticle 1Shaira CogollodoNo ratings yet
- Photosynthesis Lab HouseDocument4 pagesPhotosynthesis Lab HouseShaira CogollodoNo ratings yet
- Concept Paper Title: A Concept Paper Presented To The Senior High School Ateneo de Davao UniversityDocument6 pagesConcept Paper Title: A Concept Paper Presented To The Senior High School Ateneo de Davao UniversityShaira CogollodoNo ratings yet
- INNOSDocument3 pagesINNOSShaira CogollodoNo ratings yet
- Chapter 10 Multiple Choice Practice: CO H O O PDocument7 pagesChapter 10 Multiple Choice Practice: CO H O O PShaira CogollodoNo ratings yet
- First 1Document17 pagesFirst 1Shaira CogollodoNo ratings yet
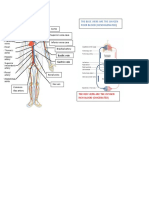
- Aorta Superior Vena Cava: The Blue Viens Are The Oxygen Poor Blood (De9OxigenatedDocument6 pagesAorta Superior Vena Cava: The Blue Viens Are The Oxygen Poor Blood (De9OxigenatedShaira CogollodoNo ratings yet
- It Group 2 Reporting FinalDocument51 pagesIt Group 2 Reporting FinalShaira CogollodoNo ratings yet
- Is FlipTop The Modern Day BalagtasanDocument18 pagesIs FlipTop The Modern Day BalagtasanShaira CogollodoNo ratings yet
- Anatomy of The Human ArmDocument2 pagesAnatomy of The Human ArmShaira CogollodoNo ratings yet
- Subject 1 (Clock-Wise) Eye Movement Observation Subject 2 (Counterclockwise) Eye Movement Observation Subject 3 (Eyes Close) Eye Movement ObservationDocument6 pagesSubject 1 (Clock-Wise) Eye Movement Observation Subject 2 (Counterclockwise) Eye Movement Observation Subject 3 (Eyes Close) Eye Movement ObservationShaira CogollodoNo ratings yet
- Plants Mineral Nutrition: Cago - Carriaga - Cogollodo - DaumarDocument46 pagesPlants Mineral Nutrition: Cago - Carriaga - Cogollodo - DaumarShaira CogollodoNo ratings yet
- Cogollodo - Plant Hormones AssignmentDocument5 pagesCogollodo - Plant Hormones AssignmentShaira CogollodoNo ratings yet
- Cogollodo - Plant Based Meal PlanDocument2 pagesCogollodo - Plant Based Meal PlanShaira CogollodoNo ratings yet
- Pical Platform Business Plan - by SlidesgoDocument94 pagesPical Platform Business Plan - by SlidesgoShaira CogollodoNo ratings yet
- Chapter 4 Basic Principles of Animal Form and FunctionDocument44 pagesChapter 4 Basic Principles of Animal Form and FunctionPrince VillacrusisNo ratings yet
- Witing TOEFL IBT Test 1Document2 pagesWiting TOEFL IBT Test 1Miriam ChiovettaNo ratings yet
- Soal Tes TOEFLDocument57 pagesSoal Tes TOEFLayu purnamaNo ratings yet
- Metabolic Pathways Study Guide MyDocument12 pagesMetabolic Pathways Study Guide Myapi-521781723No ratings yet
- Mod 8 - Kiss Notes WorksheetsDocument16 pagesMod 8 - Kiss Notes WorksheetsNeevitha JawaharNo ratings yet
- Vertebrate and Invertebrate Animal Structure NotesDocument5 pagesVertebrate and Invertebrate Animal Structure NotesJacquiline Santiago100% (1)
- Abiotic Factors in BiologyDocument20 pagesAbiotic Factors in BiologyelekviNo ratings yet
- Presentation 1Document37 pagesPresentation 1Aruji ZhukhiNo ratings yet
- TOEFL iBT - Writing SampleDocument6 pagesTOEFL iBT - Writing SampleJuan ManuelNo ratings yet
- MollesDocument32 pagesMollesSeptika RahmawatiNo ratings yet
- Homeostasis and Feedback Mechanism PDFDocument3 pagesHomeostasis and Feedback Mechanism PDFjer montillaNo ratings yet
- Complete Nutrient Composition of InsectsDocument17 pagesComplete Nutrient Composition of Insectsmbacharach8707100% (3)
- Reptiles: Section 2 Characteristics of ReptilesDocument26 pagesReptiles: Section 2 Characteristics of ReptilesNihal YildrimNo ratings yet
- Thermoregulation Article PDFDocument5 pagesThermoregulation Article PDFRio ImaniarNo ratings yet
- CK-12 Biology Chapter 19 WorksheetsDocument40 pagesCK-12 Biology Chapter 19 WorksheetsmaleskunNo ratings yet
- Fish, Amphibians, and Reptiles: Can I Find One?Document34 pagesFish, Amphibians, and Reptiles: Can I Find One?Rehan HaiderNo ratings yet
- Temperature PH and Salinity As Limiting Factors Affecting Range of ToleranceDocument4 pagesTemperature PH and Salinity As Limiting Factors Affecting Range of ToleranceCarl ReyesNo ratings yet
- Introduction To Marine Biology 4th Edition Karleskint Test BankDocument25 pagesIntroduction To Marine Biology 4th Edition Karleskint Test BankCarlosCookowmx100% (49)
- (Duellman 1990) Herpetofaunas in Neotropical Rainforests - Comparitive Composition, History, and Resource UseDocument52 pages(Duellman 1990) Herpetofaunas in Neotropical Rainforests - Comparitive Composition, History, and Resource UseDann VergaraNo ratings yet
- BT201 FINAL TERM by DINNING DISSCUSIONDocument31 pagesBT201 FINAL TERM by DINNING DISSCUSIONkabsha.raajpootNo ratings yet
- De Thi HSG Tinh Tieng Anh 12 Bac Ninh 2022-2023Document10 pagesDe Thi HSG Tinh Tieng Anh 12 Bac Ninh 2022-2023rrg rg0% (1)
- Gbio 2122 Week 1 10 by Kuya CharlesDocument18 pagesGbio 2122 Week 1 10 by Kuya CharlesChoe Yoek Soek100% (2)
- DEFINITION, EXEMPLIFICATION, CLASSIFICATION, and ADJECTIVE CLAUSESDocument16 pagesDEFINITION, EXEMPLIFICATION, CLASSIFICATION, and ADJECTIVE CLAUSESAgatha LeeNo ratings yet
- 1 - Metabolism Worksheet PDFDocument5 pages1 - Metabolism Worksheet PDFDavina SuseloNo ratings yet
- New DocumentDocument10 pagesNew DocumentIommi Arabella Paras AltreNo ratings yet
- CO1 HomeostasisDocument5 pagesCO1 HomeostasisCarmelia Jhan Fate AbrajanoNo ratings yet