Download as pdf or txt
You might also like
- SARS CoV 2andanimalCoVsDocument19 pagesSARS CoV 2andanimalCoVsmaito2270No ratings yet
- A Critique of Coronavirus (COVID-19) .Document14 pagesA Critique of Coronavirus (COVID-19) .Sujit KakadeNo ratings yet
- Breve Reseña Sobre Medicamentos y Vacunas Reutilizados para El Posible Tratamiento de COVID-19Document15 pagesBreve Reseña Sobre Medicamentos y Vacunas Reutilizados para El Posible Tratamiento de COVID-19Cristal Jewels DiamondNo ratings yet
- 2020.04.20.049924v1.full SARS-coronavirus-2 Replication in Vero E6 Cells Replication Kinetics, Rapid Adaptation and Cytopathology PDFDocument40 pages2020.04.20.049924v1.full SARS-coronavirus-2 Replication in Vero E6 Cells Replication Kinetics, Rapid Adaptation and Cytopathology PDFAngelica Ferrer TrujequeNo ratings yet
- Current Understanding of An Emerging Coronavirus UsingDocument11 pagesCurrent Understanding of An Emerging Coronavirus UsinganselmoNo ratings yet
- 2020 03 30 015008v1 Full PDFDocument25 pages2020 03 30 015008v1 Full PDFHarto EstoyNo ratings yet
- COVID MutationDocument17 pagesCOVID MutationdhairyasheelNo ratings yet
- CovDocument31 pagesCovMichelle “Ellie” TiongsonNo ratings yet
- How Nanotechnology Can Help To Combat CovidDocument23 pagesHow Nanotechnology Can Help To Combat CovidKishore PaknikarNo ratings yet
- Deep Phylogenetic-Based Clustering Analysis Uncovers New and Shared Mutations in SARS-CoV-2 Variants As A Result of Directional and Convergent EvolutionDocument14 pagesDeep Phylogenetic-Based Clustering Analysis Uncovers New and Shared Mutations in SARS-CoV-2 Variants As A Result of Directional and Convergent EvolutionDanilo RosaNo ratings yet
- Journal of Pharmaceutical Analysis: Original ArticleDocument7 pagesJournal of Pharmaceutical Analysis: Original ArticleUmmuNo ratings yet
- Dholariya2021 Article NotableAndEmergingVariantsOfSADocument8 pagesDholariya2021 Article NotableAndEmergingVariantsOfSAA.R MaheshNo ratings yet
- Evolutionary Distance of SARS-CoV-2 and Bat Coronaviruses Studied Under The NIHsupported Research Grant To EcoHealth AllianceDocument3 pagesEvolutionary Distance of SARS-CoV-2 and Bat Coronaviruses Studied Under The NIHsupported Research Grant To EcoHealth AllianceAl StatiliusNo ratings yet
- Molecular Immune Pathogenesis and Diagnosis of Covid 19 Xiaowei Li Download 2024 Full ChapterDocument29 pagesMolecular Immune Pathogenesis and Diagnosis of Covid 19 Xiaowei Li Download 2024 Full Chapterwalter.hayward549100% (11)
- Overview JournalDocument8 pagesOverview JournalDea Inthay WulanNo ratings yet
- 3985-Article Text-11543-1-10-20200510Document12 pages3985-Article Text-11543-1-10-20200510Dr. Nilesh JainNo ratings yet
- Variante SARS-CoV2 en AsintomáticosDocument12 pagesVariante SARS-CoV2 en AsintomáticosgigicarvajalNo ratings yet
- Genomic Characterization of The 2019 Novel Human-Pathogenic Coronavirus Isolated From A Patient With Atypical Pneumonia After Visiting WuhanDocument18 pagesGenomic Characterization of The 2019 Novel Human-Pathogenic Coronavirus Isolated From A Patient With Atypical Pneumonia After Visiting WuhanMuhamad vero SyatiaNo ratings yet
- COVID-19 Gain of FunctionDocument5 pagesCOVID-19 Gain of FunctionRed Cloud ShamanNo ratings yet
- Rañola Micro-Para Covid-19 PrelimsDocument6 pagesRañola Micro-Para Covid-19 PrelimsAndrey Mary RanolaNo ratings yet
- Articulo CoronavirusDocument50 pagesArticulo CoronavirusLAURA MARCELA BARRENECHE CALLENo ratings yet
- Omeprazole in Covid 19 Germany StudyDocument28 pagesOmeprazole in Covid 19 Germany StudyTanushree PanjariNo ratings yet
- In Silico Evaluation of Flavonoids As Effective Antiviral Agents On The SpikeDocument12 pagesIn Silico Evaluation of Flavonoids As Effective Antiviral Agents On The SpikeLuis MartinezNo ratings yet
- COVID-19 Coronavirus Spike Protein Analysis FDocument66 pagesCOVID-19 Coronavirus Spike Protein Analysis Fasdjjjjss100% (1)
- CUREUSCORONAREVIEWDocument14 pagesCUREUSCORONAREVIEWummu yayaNo ratings yet
- Wetransfer - ProjectDocument40 pagesWetransfer - ProjectĐinh ThắngNo ratings yet
- Mechanisms of Sars-Cov-2 Transmission and Pathogenesis: ReviewDocument16 pagesMechanisms of Sars-Cov-2 Transmission and Pathogenesis: ReviewRocio GmarNo ratings yet
- Bioactive Natural Compounds Against Human Coronavirus - 2020 - Acta PharmaceuticDocument12 pagesBioactive Natural Compounds Against Human Coronavirus - 2020 - Acta PharmaceuticMohammed Shuaib AhmedNo ratings yet
- Antivirals Against Coronaviruses: Candidate Drugs For Sars-Cov-2 Treatment?Document23 pagesAntivirals Against Coronaviruses: Candidate Drugs For Sars-Cov-2 Treatment?zizaviquisaNo ratings yet
- Bioorganic Chemistry: MD Jawaid Akhtar TDocument9 pagesBioorganic Chemistry: MD Jawaid Akhtar TRetno Tri RahayuNo ratings yet
- Structural Proteins in Severe Acute Respiratory Syndrome Coronavirus-2Document10 pagesStructural Proteins in Severe Acute Respiratory Syndrome Coronavirus-2automationenggNo ratings yet
- Bat Origin of Human CoronavirusesDocument10 pagesBat Origin of Human CoronavirusesThe Truth ChroniclesNo ratings yet
- 2005 ArtiDocument8 pages2005 ArtiasdjjjjssNo ratings yet
- Microorganisms: Sars-Cov-2 Pandemic: Not The First, Not The LastDocument10 pagesMicroorganisms: Sars-Cov-2 Pandemic: Not The First, Not The LastDeepayancNo ratings yet
- Project 1Document64 pagesProject 1haleem ullah khanNo ratings yet
- Innate and Adaptive Immune Responses Against CoronavirusDocument17 pagesInnate and Adaptive Immune Responses Against CoronavirusHARDITA SALSABILA FORTUNA HARDITA SALSABILA FORTUNANo ratings yet
- Viruses 14 02194 v3 1Document28 pagesViruses 14 02194 v3 1marab77No ratings yet
- An Insight Into The Pathogenesis, Diagnosis and Treatment of SARS-Cov-2 With A Synopsis of Situation in Malaysia During The 2020 OutbreakDocument12 pagesAn Insight Into The Pathogenesis, Diagnosis and Treatment of SARS-Cov-2 With A Synopsis of Situation in Malaysia During The 2020 OutbreakUMYU Journal of Microbiology Research (UJMR)No ratings yet
- Bioinformatics Analysis of The Recent MERS-CoV With Special Reference To The Virus-Encoded Spike ProteinDocument10 pagesBioinformatics Analysis of The Recent MERS-CoV With Special Reference To The Virus-Encoded Spike ProteinInternational Medical PublisherNo ratings yet
- Pathogens: Biology and Pathogenesis of Sars-Cov-2: Understandings For Therapeutic Developments Against Covid-19Document27 pagesPathogens: Biology and Pathogenesis of Sars-Cov-2: Understandings For Therapeutic Developments Against Covid-19neftaliNo ratings yet
- Covid-19 - A Pandemic by Sars-Cov-2: Harsh R Darji (J18Imt601)Document3 pagesCovid-19 - A Pandemic by Sars-Cov-2: Harsh R Darji (J18Imt601)Harsh Darji100% (1)
- 1 s2.0 S1567134820303336 MainDocument15 pages1 s2.0 S1567134820303336 MainKaren Lisseth Jimenez ArmijosNo ratings yet
- Predicting Commercially Available Antiviral Drugs Tha - 2020 - Computational andDocument7 pagesPredicting Commercially Available Antiviral Drugs Tha - 2020 - Computational andOksana BagreevaNo ratings yet
- Diagnosing Covid 19 by CRISPR TechnologyDocument5 pagesDiagnosing Covid 19 by CRISPR TechnologyEditor IJTSRDNo ratings yet
- Ad26.Cov2.S Prevents Sars-Cov-2 Induced Pathways of Inflammation and Thrombosis in HamstersDocument46 pagesAd26.Cov2.S Prevents Sars-Cov-2 Induced Pathways of Inflammation and Thrombosis in Hamsterscristianismo sem MáscaraNo ratings yet
- Project 1 (1 100 Page)Document47 pagesProject 1 (1 100 Page)haleem ullah khanNo ratings yet
- 2020.01.30.926477v1.full - PDF Evolucion Estimada Del 2019nCoVDocument17 pages2020.01.30.926477v1.full - PDF Evolucion Estimada Del 2019nCoVRENE RIVASNo ratings yet
- CoronavirusDocument12 pagesCoronavirusZubair GhouriNo ratings yet
- J Antiviral 2020 104742Document20 pagesJ Antiviral 2020 104742hanzNo ratings yet
- European Journal of Pharmacology: Gayathri Krishna, Vinod Soman Pillai, Mohanan Valiya VeettilDocument11 pagesEuropean Journal of Pharmacology: Gayathri Krishna, Vinod Soman Pillai, Mohanan Valiya VeettilautomationenggNo ratings yet
- Viruses: Canine Enteric Coronaviruses: Emerging Viral Pathogens With Distinct Recombinant Spike ProteinsDocument14 pagesViruses: Canine Enteric Coronaviruses: Emerging Viral Pathogens With Distinct Recombinant Spike ProteinsYovan BlancoNo ratings yet
- Preprints202002 0308 v1 PDFDocument16 pagesPreprints202002 0308 v1 PDFCarlos Andres Padilla JaramilloNo ratings yet
- The Roles of Signaling Pathways in Sars Cov 2 Infection Lessons Learned From Sars Cov and Mers CovDocument22 pagesThe Roles of Signaling Pathways in Sars Cov 2 Infection Lessons Learned From Sars Cov and Mers CovTufik NaderNo ratings yet
- Covide 19 BeginigDocument14 pagesCovide 19 BeginigHamza MehalliNo ratings yet
- The Taxonomy Host Range and Pathogenicity of CoronDocument28 pagesThe Taxonomy Host Range and Pathogenicity of CoronAbdul Latif Nur MuizNo ratings yet
- Coronavirus: COVID-19 Pandemic Coronavirus Disease 2019 Severe Acute Respiratory Syndrome Coronavirus 2Document26 pagesCoronavirus: COVID-19 Pandemic Coronavirus Disease 2019 Severe Acute Respiratory Syndrome Coronavirus 23esNo ratings yet
- Benzalkonium CL Rinsed VirusDocument12 pagesBenzalkonium CL Rinsed VirusGeneral Division MedicineNo ratings yet
- Clinical and Molecular Relationships Between COVID-19 and Feline Infectious Peritonitis (FIP)Document23 pagesClinical and Molecular Relationships Between COVID-19 and Feline Infectious Peritonitis (FIP)Reza Alfitra MutiaraNo ratings yet
- Biology of Sars-Cov-2Document4 pagesBiology of Sars-Cov-2J. L.No ratings yet
- Overwhelming Mutations or SNPs of SARS-CoV-2: A Point of CautionDocument4 pagesOverwhelming Mutations or SNPs of SARS-CoV-2: A Point of CautionDanilo RosaNo ratings yet
- Quasispecies of SARS-CoV-2 Revealed by Single Nucleotide Polymorphisms (SNPS) AnalysisDocument18 pagesQuasispecies of SARS-CoV-2 Revealed by Single Nucleotide Polymorphisms (SNPS) AnalysisDanilo RosaNo ratings yet
- Deep Phylogenetic-Based Clustering Analysis Uncovers New and Shared Mutations in SARS-CoV-2 Variants As A Result of Directional and Convergent EvolutionDocument14 pagesDeep Phylogenetic-Based Clustering Analysis Uncovers New and Shared Mutations in SARS-CoV-2 Variants As A Result of Directional and Convergent EvolutionDanilo RosaNo ratings yet
- Factors Associated With Covid-19-Related Death Using OpensafelyDocument17 pagesFactors Associated With Covid-19-Related Death Using OpensafelyDanilo RosaNo ratings yet
- C Elegans Enriquecimento BioinfoDocument10 pagesC Elegans Enriquecimento BioinfoDanilo RosaNo ratings yet
- Wou 1408 ApprovedDocument13 pagesWou 1408 ApprovedDanilo RosaNo ratings yet
- Nutrition in Plants Edexcel IGCSEDocument7 pagesNutrition in Plants Edexcel IGCSETZ AZNo ratings yet
- NSEC Solved Paper 2012Document19 pagesNSEC Solved Paper 2012swapnil narayanNo ratings yet
- Annotated Source List: Main TopicDocument6 pagesAnnotated Source List: Main Topicapi-336220957No ratings yet
- Integral City 2.0 Online Conference Proceedings: Executive SummaryDocument3 pagesIntegral City 2.0 Online Conference Proceedings: Executive SummaryMarilyn HamiltonNo ratings yet
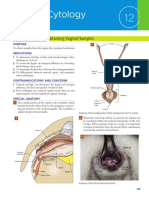
- Vaginal CytologyDocument4 pagesVaginal Cytologyابراهيم عبدالله100% (1)
- Sbi 3u1 Ap Bio Practice ExamDocument27 pagesSbi 3u1 Ap Bio Practice ExamasjiodjwNo ratings yet
- Aestivation: Latin American English Dormancy Hibernation MetabolicDocument3 pagesAestivation: Latin American English Dormancy Hibernation MetabolicMalikNoumanAwanNo ratings yet
- Cilia 2018Document1 pageCilia 2018Anna Beatriz Silva EspindolaNo ratings yet
- Philsci Questionnaire FinalDocument15 pagesPhilsci Questionnaire Finalandrea dyanne AzoresNo ratings yet
- Factors Affecting Starch DigestionDocument2 pagesFactors Affecting Starch DigestionNikki Ticman100% (1)
- BPak's MI40-X - CEP Training BlueprintDocument22 pagesBPak's MI40-X - CEP Training BlueprintJose Rincon75% (8)
- Biomarkers of PDDocument14 pagesBiomarkers of PDAmey MoreNo ratings yet
- AI & ALIENS, The Truthful Analysis - Jeremy GriffithDocument34 pagesAI & ALIENS, The Truthful Analysis - Jeremy Griffithvas_el_ionNo ratings yet
- Aplikasi Teknologi Budidaya Kelengkeng Super Sleman Di Padukuhan GejayanDocument5 pagesAplikasi Teknologi Budidaya Kelengkeng Super Sleman Di Padukuhan GejayanMelly AntikaNo ratings yet
- Blotting TechniquesDocument19 pagesBlotting TechniquesGhilli TecNo ratings yet
- Ivan Illich Medical NemesisDocument332 pagesIvan Illich Medical NemesisRăzvan Necula100% (2)
- A.ANBU M.SC.,B.ED., PH: 8508429396 Vivek Vidyalaya Matric Higher Secondary SchoolDocument6 pagesA.ANBU M.SC.,B.ED., PH: 8508429396 Vivek Vidyalaya Matric Higher Secondary SchoolSilva scary svNo ratings yet
- Biopharmaceutics Unit 4Document19 pagesBiopharmaceutics Unit 4Niki Rokade DahaleNo ratings yet
- JHG 201236Document3 pagesJHG 201236Iva TintiloNo ratings yet
- Plastination-WPS OfficeDocument3 pagesPlastination-WPS OfficeAmina RahimNo ratings yet
- Limbs in Whales and Limblessness in Other VertebratesDocument14 pagesLimbs in Whales and Limblessness in Other VertebratesJay LeeNo ratings yet
- Kuliah Rickets PBLDocument43 pagesKuliah Rickets PBLNovasuryatiNo ratings yet
- Dls 213 - Selected Topics in General Biology: University of Northern PhilippinesDocument28 pagesDls 213 - Selected Topics in General Biology: University of Northern PhilippinesKennedy Fieldad VagayNo ratings yet
- Cleaning Sorting GradingDocument17 pagesCleaning Sorting GradingMuhammad HussainNo ratings yet
- Webinar EU Medical Device Regulation Jun 28 2017 v2Document42 pagesWebinar EU Medical Device Regulation Jun 28 2017 v2Thulasi VasudevanNo ratings yet
- HistologyDocument63 pagesHistologyMichelle ThereseNo ratings yet
- Cell Lab ReportDocument3 pagesCell Lab ReportJosé SuárezNo ratings yet
- Logical Framework Analysis As A Tool For Tropical Fisheries ManagementDocument11 pagesLogical Framework Analysis As A Tool For Tropical Fisheries ManagementPeter A Murray100% (2)
- Article 1 - Nantechonology-1Document19 pagesArticle 1 - Nantechonology-1Alexander LuNo ratings yet
- Tongkat AliDocument3 pagesTongkat AliRifa Rania RinaWanNo ratings yet