Download as doc, pdf, or txt
You might also like
- Goodness of God Key of EDocument2 pagesGoodness of God Key of Estephen2buizonNo ratings yet
- Lecture 7 Population Properties and Dynamics BWDocument69 pagesLecture 7 Population Properties and Dynamics BWPeter JimenezNo ratings yet
- Chapter 2 Envi Sci 1Document32 pagesChapter 2 Envi Sci 1ANDREW ESLOPOR ESPINOSANo ratings yet
- Population DynamicsDocument5 pagesPopulation DynamicsUmair100% (1)
- Population Dynamics Study by FahmidDocument9 pagesPopulation Dynamics Study by FahmidFahmid Tousif KhanNo ratings yet
- 12 Population BiologyDocument22 pages12 Population Biologyजलन्धरNo ratings yet
- Birth Death ChangesDocument5 pagesBirth Death ChangesJane OrsalNo ratings yet
- Chapter 1 - PopulationDocument41 pagesChapter 1 - PopulationOrlando SaliviaNo ratings yet
- Biology For All 303 Single Species DynamicsDocument9 pagesBiology For All 303 Single Species DynamicswealthiykejephNo ratings yet
- Unit 4 Ecosystems: Stability and ChangeDocument8 pagesUnit 4 Ecosystems: Stability and ChangeKennade BarberNo ratings yet
- Population CharacteristicsDocument14 pagesPopulation CharacteristicsJulieann Salvacion OberesNo ratings yet
- Population CharacteristicsDocument14 pagesPopulation CharacteristicsJulieann Salvacion OberesNo ratings yet
- Lecture 10: Population Growth: Dafeng Hui Office: Harned Hall 320 Phone: 963-5777 Email: Dhui@tnstate - EduDocument41 pagesLecture 10: Population Growth: Dafeng Hui Office: Harned Hall 320 Phone: 963-5777 Email: Dhui@tnstate - Eduputt510No ratings yet
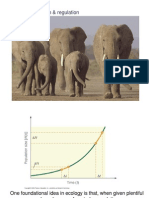
- Population Growth & RegulationDocument23 pagesPopulation Growth & RegulationJong FriasNo ratings yet
- Population Is A Group of Organisms of The Same Species Which Occupies The Same Habitat and Forms A Single Functional Unit of Biotic CommunityDocument5 pagesPopulation Is A Group of Organisms of The Same Species Which Occupies The Same Habitat and Forms A Single Functional Unit of Biotic CommunityHanelyn HenryNo ratings yet
- Characteristics of PopulationDocument9 pagesCharacteristics of PopulationmosesmahamudukamwendoNo ratings yet
- Quantitative EcologyDocument123 pagesQuantitative EcologyKrishnan JeevanNo ratings yet
- Population Growth - Intraspecific CompetitionDocument4 pagesPopulation Growth - Intraspecific Competitionmuslim united 2018No ratings yet
- 4apopulation DynamicsDocument98 pages4apopulation DynamicsJerome FlorendoNo ratings yet
- Population Biology ConceptDocument18 pagesPopulation Biology ConceptGilmar BaunNo ratings yet
- Population EcologyDocument41 pagesPopulation Ecologyshahrafiq79No ratings yet
- Lesson 3 - Population EcologyDocument10 pagesLesson 3 - Population EcologyAdriel MarasiganNo ratings yet
- Human Population and The EnvironmentDocument37 pagesHuman Population and The EnvironmentlibrarianNo ratings yet
- APSEd Population Ecology Self-Read Material Lyst1402Document8 pagesAPSEd Population Ecology Self-Read Material Lyst1402Shubham NaikNo ratings yet
- Ge Elec 1Document12 pagesGe Elec 1justnothing010100No ratings yet
- Core Areas of Environmental ScienceDocument6 pagesCore Areas of Environmental ScienceKIM IVAN SUMAGAYSAYNo ratings yet
- Economic Policies in India 2Document16 pagesEconomic Policies in India 2itantenna1No ratings yet
- Logistic Function: For The Recurrence Relation, SeeDocument8 pagesLogistic Function: For The Recurrence Relation, SeeMazhar ZamanNo ratings yet
- Introduction To Ecology: Define The Population byDocument6 pagesIntroduction To Ecology: Define The Population byLim Yi FeiNo ratings yet
- GEC 21 PopulationNotesDocument5 pagesGEC 21 PopulationNotesMika TrincheraNo ratings yet
- Organisms and Populations - PMDDocument15 pagesOrganisms and Populations - PMDRajika Sri.VNo ratings yet
- Population Ecology - ReproductionDocument2 pagesPopulation Ecology - ReproductionEJNo ratings yet
- PopulationDocument15 pagesPopulationBarshaNo ratings yet
- Lecture Population EcologyDocument75 pagesLecture Population Ecologyrkv100% (3)
- Population EcologyDocument15 pagesPopulation EcologyFarman UddinNo ratings yet
- Population Ecology: Aecc-I +3 1 YearDocument32 pagesPopulation Ecology: Aecc-I +3 1 YearAnita kumari SahuNo ratings yet
- Population Ecology, DemecologyDocument30 pagesPopulation Ecology, DemecologyNazarbek Auyeskhan100% (1)
- Human Population UpdatedDocument53 pagesHuman Population UpdatedXeyl NarvazaNo ratings yet
- Population: World Population Through Ages PopulationDocument8 pagesPopulation: World Population Through Ages PopulationShagnik RoyNo ratings yet
- FFB-311-L3,4 & 6 (Popl - GCurve)Document16 pagesFFB-311-L3,4 & 6 (Popl - GCurve)joshigautaNo ratings yet
- Unit SixDocument53 pagesUnit Sixtesfamlaksolomon33No ratings yet
- Ecology of Populations: PopulationDocument23 pagesEcology of Populations: PopulationJÜnn BatacNo ratings yet
- Montero, Lina Riza N. BS Bio IVDocument3 pagesMontero, Lina Riza N. BS Bio IVMIK206No ratings yet
- Variation (Continuous/ Discontinuous)Document6 pagesVariation (Continuous/ Discontinuous)Immanuel LashleyNo ratings yet
- Biology Chapter 5 PopulationsDocument52 pagesBiology Chapter 5 PopulationsLesleyNo ratings yet
- Zge 4301-Population EcologyDocument45 pagesZge 4301-Population Ecologymikeevillar97No ratings yet
- GE 10 Environmental Science: Prepared By: Marygrace P. Sanopal, LPTDocument17 pagesGE 10 Environmental Science: Prepared By: Marygrace P. Sanopal, LPTVERGIE LAPINIGNo ratings yet
- Growth Is Likely To Refer To Any Known Organism, But This Article Deals Mostly With The Application of The Term ToDocument4 pagesGrowth Is Likely To Refer To Any Known Organism, But This Article Deals Mostly With The Application of The Term ToshielamariNo ratings yet
- Population Growth Bread Mould StudentDocument7 pagesPopulation Growth Bread Mould StudentJohn OsborneNo ratings yet
- Bio 307 General Ecology IIIDocument26 pagesBio 307 General Ecology IIIokonkwojohnchuks1100% (1)
- What Is Demography?: (CITATION WGA16 /L 1033)Document10 pagesWhat Is Demography?: (CITATION WGA16 /L 1033)Coma Mary Shane O.No ratings yet
- Population 2: Ekologi TumbuhanDocument32 pagesPopulation 2: Ekologi TumbuhanDitaMenonNo ratings yet
- Properties of PopulationDocument26 pagesProperties of PopulationJannat FirdousNo ratings yet
- DEMOGRAPHYDocument7 pagesDEMOGRAPHYJeyanthiNo ratings yet
- Chapter 2: Population Ecology: Return To Table of ContentsDocument11 pagesChapter 2: Population Ecology: Return To Table of ContentsPiolo Mar RomeroNo ratings yet
- Density Independent and Depedent Study PDFDocument12 pagesDensity Independent and Depedent Study PDFKrystal An RemoladoNo ratings yet
- The Uplift Project: Enhancing and Propagating Intelligence and LongevityFrom EverandThe Uplift Project: Enhancing and Propagating Intelligence and LongevityNo ratings yet
- Preclinical Behavioral Science and Social Sciences Review 2023: For USMLE Step 1 and COMLEX-USA Level 1From EverandPreclinical Behavioral Science and Social Sciences Review 2023: For USMLE Step 1 and COMLEX-USA Level 1No ratings yet
- Euthenics, the science of controllable environment: A plea for better living conditions as a first step toward higher human efficiencyFrom EverandEuthenics, the science of controllable environment: A plea for better living conditions as a first step toward higher human efficiencyNo ratings yet
- Sample Diary Curriculum Map SUBJECT: Mathematics QUARTER: Second Grade Level: 10 TOPIC: CircleDocument3 pagesSample Diary Curriculum Map SUBJECT: Mathematics QUARTER: Second Grade Level: 10 TOPIC: CircleRichimon LicerioNo ratings yet
- Cassava 2Document5 pagesCassava 2CESAR JAMESNo ratings yet
- Painting Still Water in WatercolorDocument4 pagesPainting Still Water in WatercolorRuiNo ratings yet
- Zikr-e-Masoom (A.s.)Document262 pagesZikr-e-Masoom (A.s.)Shian-e-Ali Network0% (1)
- A Term Paper On Justice Holmes'S Concept of Law: Adjunct Faculty Mr. Dev Mahat Nepal Law CampusDocument4 pagesA Term Paper On Justice Holmes'S Concept of Law: Adjunct Faculty Mr. Dev Mahat Nepal Law Campusrahul jhaNo ratings yet
- China Pakistan Economic Corridor CPEC IsDocument12 pagesChina Pakistan Economic Corridor CPEC IsNasir HussainNo ratings yet
- From Lovemaking To Superconsciousness-0shoDocument31 pagesFrom Lovemaking To Superconsciousness-0shoRAGUPATHYNo ratings yet
- III Year A EEEDocument2 pagesIII Year A EEEshenbagaraman cseNo ratings yet
- The Learners Demonstrate An Understanding of The Significance of Studying Culture, Society, and PoliticsDocument2 pagesThe Learners Demonstrate An Understanding of The Significance of Studying Culture, Society, and PoliticsMabelle BagtasosNo ratings yet
- Air Is Allowed To Flow From A Large Reservoir ThroDocument3 pagesAir Is Allowed To Flow From A Large Reservoir Throsid202pkNo ratings yet
- Writing Strategy and Tips - CAE PDFDocument11 pagesWriting Strategy and Tips - CAE PDFAntoSeitanNo ratings yet
- Course Title Understanding Culture, Society, & Politics Instructor Mieca Aguinaldo Course Code Course DescriptionDocument6 pagesCourse Title Understanding Culture, Society, & Politics Instructor Mieca Aguinaldo Course Code Course DescriptionSJHC 21No ratings yet
- It8078 Web Design and ManagementDocument27 pagesIt8078 Web Design and ManagementsanjayNo ratings yet
- FIN201 CF T3 2021 BBUS Unit Guide V2 07092021Document11 pagesFIN201 CF T3 2021 BBUS Unit Guide V2 07092021Nguyen Quynh AnhNo ratings yet
- Rickett'S: Visual Treatment Objective (V.T.O)Document4 pagesRickett'S: Visual Treatment Objective (V.T.O)MariyamNo ratings yet
- Man-In-Simulant Test (MIST) For Protective EnsemblesDocument11 pagesMan-In-Simulant Test (MIST) For Protective EnsemblesAhmad Zubair RasulyNo ratings yet
- RKSV Securities India Private Limited Registered Corporate AddressDocument2 pagesRKSV Securities India Private Limited Registered Corporate AddressAjarihant OrthoNo ratings yet
- SOLO MATH LESSON PLAN - G10 - Ccluster 5Document6 pagesSOLO MATH LESSON PLAN - G10 - Ccluster 5April Rose Caquilala GalauraNo ratings yet
- Confidence: A Pentecostal Paper For Great Britain April-June 1922 No. 129.Document16 pagesConfidence: A Pentecostal Paper For Great Britain April-June 1922 No. 129.James MayugaNo ratings yet
- What Is The Difference Between Graduate and UndergraduateDocument6 pagesWhat Is The Difference Between Graduate and UndergraduateWinnie OchiengNo ratings yet
- FinAcc MCQ Sem2 PDFDocument11 pagesFinAcc MCQ Sem2 PDFBhavesh Rathod100% (2)
- Diet PlanDocument4 pagesDiet PlanRahul BhattacharjeeNo ratings yet
- Final ExamDocument3 pagesFinal ExamFaidah PangandamanNo ratings yet
- Startup Law OverviewDocument11 pagesStartup Law OverviewAgibise AkanliseNo ratings yet
- FINAL NML-NEW ADVT 02by2020 PDFDocument9 pagesFINAL NML-NEW ADVT 02by2020 PDFsiddharth singhNo ratings yet
- Role of Agnikarma in Pain ManagementDocument9 pagesRole of Agnikarma in Pain ManagementPoonam KailoriaNo ratings yet
- Fy2023 FWS Budget RequestDocument8 pagesFy2023 FWS Budget RequestinforumdocsNo ratings yet
- Vampires Werewolves and Witches Oh MyDocument51 pagesVampires Werewolves and Witches Oh Myangelg29850% (1)
- The Literature of Bibliometrics Scientometrics and Informetrics-2Document24 pagesThe Literature of Bibliometrics Scientometrics and Informetrics-2Juan Ruiz-UrquijoNo ratings yet