Download as pdf or txt
You might also like
- Research ProposalDocument2 pagesResearch Proposalapi-252462660100% (1)
- Partogram Professor Adel Abul-Heija FRCOGDocument45 pagesPartogram Professor Adel Abul-Heija FRCOGNizar Rabadi100% (2)
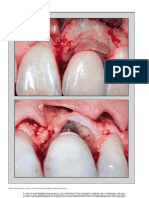
- The International Journal of Periodontics & Restorative DentistryDocument8 pagesThe International Journal of Periodontics & Restorative DentistryjonvargasNo ratings yet
- 01 Immunonutrition Support For AthletesDocument4 pages01 Immunonutrition Support For AthletesAdid PunyaNo ratings yet
- Surgery 00aDocument8 pagesSurgery 00aamelindavaNo ratings yet
- Capitulo 5 Gut MicrobiotaDocument19 pagesCapitulo 5 Gut MicrobiotaFco. Javier Grijota PérezNo ratings yet
- Xie Et Al 2023 A Major Mechanism For ImmunomodulationDocument13 pagesXie Et Al 2023 A Major Mechanism For Immunomodulationtaha242004No ratings yet
- Immunonutrition in Gastric Cancer Surgical PatientsDocument2 pagesImmunonutrition in Gastric Cancer Surgical Patientsusuario101122No ratings yet
- As Stated Previously, in A Few Hours of Exercise or Work in The Heat, Water Loss orDocument10 pagesAs Stated Previously, in A Few Hours of Exercise or Work in The Heat, Water Loss orEric Mayo Dagradi Kabat100% (1)
- Examining The Interaction Between Exercise, Gut Microbiota, and NeurodegenerationDocument17 pagesExamining The Interaction Between Exercise, Gut Microbiota, and NeurodegenerationgcabanNo ratings yet
- Denou (2016) - Gut Microbiota & ExerciseDocument12 pagesDenou (2016) - Gut Microbiota & ExerciselchitkayNo ratings yet
- 2012 The Impact of Probiotics andDocument7 pages2012 The Impact of Probiotics andAleivi PérezNo ratings yet
- Desordenes Gastrointestinales PDFDocument7 pagesDesordenes Gastrointestinales PDFMvz Mary Cielo Fernández FuentesNo ratings yet
- Ramadhan Et CancerDocument12 pagesRamadhan Et CancerSarah MellaliNo ratings yet
- Immune Adaptation To Chronic Intense Exercise Training: New Microarray EvidenceDocument10 pagesImmune Adaptation To Chronic Intense Exercise Training: New Microarray EvidenceTiago MachadoNo ratings yet
- tmp7DE6 TMPDocument5 pagestmp7DE6 TMPFrontiersNo ratings yet
- tmp38C0 TMPDocument5 pagestmp38C0 TMPFrontiersNo ratings yet
- tmp7AC5 TMPDocument5 pagestmp7AC5 TMPFrontiersNo ratings yet
- Spirulina ImuneDocument12 pagesSpirulina ImuneBruna Do CarmoNo ratings yet
- Mecanismo Dos ProbioticosDocument10 pagesMecanismo Dos ProbioticosFernanda ToledoNo ratings yet
- Beyond The Genes: Unveiling The Power of Metabolome Interactions For Biomedical ProgressDocument4 pagesBeyond The Genes: Unveiling The Power of Metabolome Interactions For Biomedical ProgressIJAR JOURNALNo ratings yet
- Microbiota en El Deprte 2021Document15 pagesMicrobiota en El Deprte 2021RosaNo ratings yet
- The Role of Ganoderma Lucidum in Ulcerative Colitis Revision 3 Submisi JgheDocument10 pagesThe Role of Ganoderma Lucidum in Ulcerative Colitis Revision 3 Submisi JgheShabrina MaharaniNo ratings yet
- Role of Plant Based Functional Food in ImmunologyDocument10 pagesRole of Plant Based Functional Food in ImmunologyelsayedmohammedmaalyNo ratings yet
- Molecules 28 06811Document35 pagesMolecules 28 06811Nejc KovačNo ratings yet
- Campos 2018Document17 pagesCampos 2018ngvy520201No ratings yet
- Pumpa 2019Document6 pagesPumpa 2019dyahpuspardaniNo ratings yet
- Diet and Nutrients and ImmuneDocument21 pagesDiet and Nutrients and ImmuneKaty Carreño RojasNo ratings yet
- Ncomms12015 Scleroza Multipla PDFDocument11 pagesNcomms12015 Scleroza Multipla PDFLUTICHIEVICI NATALIANo ratings yet
- Biopolym - Cell 2020 36 1 022 enDocument13 pagesBiopolym - Cell 2020 36 1 022 enАнна ШаповаловаNo ratings yet
- Review Article: Exercise Modifies The Gut Microbiota With Positive Health EffectsDocument8 pagesReview Article: Exercise Modifies The Gut Microbiota With Positive Health EffectsVanNo ratings yet
- Effect of High-Resistance Training at Intestinal Level - UAMDocument11 pagesEffect of High-Resistance Training at Intestinal Level - UAMJULIETANo ratings yet
- The Athletic Gut MicrobiotaDocument33 pagesThe Athletic Gut MicrobiotaEmanuel Roca CastroNo ratings yet
- Japplphysiol 00277 2018Document11 pagesJapplphysiol 00277 2018Misael VargasNo ratings yet
- Using Probioticos in Gastrointestinal DisordersDocument7 pagesUsing Probioticos in Gastrointestinal DisordersRonald CampoverdeNo ratings yet
- 1 s2.0 S0002916522004385 MainDocument11 pages1 s2.0 S0002916522004385 Maincarlos santamariaNo ratings yet
- DANTAS 2020 Exercise-Induced Increases in Insulin Sensitivity After Bariatric Surgery Are Mediated by Muscle Extracellular Matrix RemodelingDocument17 pagesDANTAS 2020 Exercise-Induced Increases in Insulin Sensitivity After Bariatric Surgery Are Mediated by Muscle Extracellular Matrix RemodelingCris AquinoNo ratings yet
- Present As IDocument9 pagesPresent As Iaini rahmawatiNo ratings yet
- The Effects of Twenty-Four Nutrients and Phytonutrients On Immune System Function and Inflammation A Narrative ReviewDocument44 pagesThe Effects of Twenty-Four Nutrients and Phytonutrients On Immune System Function and Inflammation A Narrative ReviewVeronica Ferreiro BlazeticNo ratings yet
- A Diet of U S Military Food Rations Alters Gut Micro 2019 The Journal of NuDocument11 pagesA Diet of U S Military Food Rations Alters Gut Micro 2019 The Journal of NuhngNo ratings yet
- Microbiota-Gut-Brain Axis: Modulator of Host Metabolism and AppetiteDocument19 pagesMicrobiota-Gut-Brain Axis: Modulator of Host Metabolism and AppetiterizkiNo ratings yet
- Five Tips To Help Cultivate A Healthy MicrobiomeDocument9 pagesFive Tips To Help Cultivate A Healthy MicrobiomeCristina Дима100% (1)
- Aceite de Semilla de Granada en EjercicioDocument9 pagesAceite de Semilla de Granada en EjercicioHiramGzhizmiguyNo ratings yet
- Quercetin FactsheetDocument3 pagesQuercetin FactsheetFelipe BarileNo ratings yet
- The Combination of Sport and Sport-Specific Diet Is Associated With Characteristics of Gut Microbiota: An Observational StudyDocument10 pagesThe Combination of Sport and Sport-Specific Diet Is Associated With Characteristics of Gut Microbiota: An Observational StudyEdgar SánchezNo ratings yet
- Artigo Sobre Probióticos No EsporteDocument9 pagesArtigo Sobre Probióticos No Esporterenato cruzNo ratings yet
- Ibrahim Et Al., 2016 PDFDocument11 pagesIbrahim Et Al., 2016 PDFDiba Eka DiputriNo ratings yet
- Potential Role of Nutrients On Immunity: Mini ReviewDocument11 pagesPotential Role of Nutrients On Immunity: Mini ReviewEvi NopiantiiNo ratings yet
- Artigo Eixo Intestino e MusculoDocument19 pagesArtigo Eixo Intestino e MusculoAgenda da TaiseNo ratings yet
- Immunomodulatory Effect of Fibres, Probiotics and Synbiotics in Different Life-StagesDocument9 pagesImmunomodulatory Effect of Fibres, Probiotics and Synbiotics in Different Life-StagesAdán Josué Cetina DorantesNo ratings yet
- Presentation Gut Brain August 2018Document17 pagesPresentation Gut Brain August 2018Sribharathi Indrajith100% (1)
- Artigo Pics 4 PDFDocument9 pagesArtigo Pics 4 PDFGabriela Brito de Lima SilvaNo ratings yet
- Effects of Microbiota-Targetted Treatments (Probiotik, Probiotik Food, and Prebiotik) On Inflammatory Bowel DiseaseDocument12 pagesEffects of Microbiota-Targetted Treatments (Probiotik, Probiotik Food, and Prebiotik) On Inflammatory Bowel DiseaseBagas FairusyahNo ratings yet
- Gillies 2003Document6 pagesGillies 2003puspitaNo ratings yet
- Nutrients 14 03985 v2Document17 pagesNutrients 14 03985 v2Lisa GonçalvesNo ratings yet
- Exercise, Nutrition, and Immune Function: Journal of Sports Sciences February 2004Document12 pagesExercise, Nutrition, and Immune Function: Journal of Sports Sciences February 2004Federico CilloNo ratings yet
- Gut Brain MicrobiotaDocument19 pagesGut Brain MicrobiotaOana CornutiuNo ratings yet
- Role of The Gut Microbiota in Defining Human HealthDocument15 pagesRole of The Gut Microbiota in Defining Human HealthVictor ResendizNo ratings yet
- Microbiota IntestinalDocument48 pagesMicrobiota IntestinalDina OliveiraNo ratings yet
- Sistema InmuneDocument40 pagesSistema InmuneLalo Rmz17No ratings yet
- Fibras e MicrobiotaDocument10 pagesFibras e Microbiotataelesoliveira.tNo ratings yet
- Cultivating Wellness The Comprehensive Guide to Probiotics and Human HealthFrom EverandCultivating Wellness The Comprehensive Guide to Probiotics and Human HealthNo ratings yet
- Total Scar Management: From Lasers to Surgery for Scars, Keloids, and Scar ContracturesFrom EverandTotal Scar Management: From Lasers to Surgery for Scars, Keloids, and Scar ContracturesRei OgawaNo ratings yet
- Bereavement Leave PolicyDocument2 pagesBereavement Leave PolicyAshley PeartNo ratings yet
- Speed & Agility: Practical SessionDocument7 pagesSpeed & Agility: Practical SessionAshley PeartNo ratings yet
- Circuit Training On The Handball Court-Ehf MasterDocument45 pagesCircuit Training On The Handball Court-Ehf MasterAshley PeartNo ratings yet
- 2021 WNT Selection Procedures - Final DraftDocument11 pages2021 WNT Selection Procedures - Final DraftAshley PeartNo ratings yet
- Competitive Demands of Elite Handball: April 2014Document9 pagesCompetitive Demands of Elite Handball: April 2014Ashley PeartNo ratings yet
- Close Contact Who Are Fully VaccinatedDocument3 pagesClose Contact Who Are Fully VaccinatedMark Anthony EstacioNo ratings yet
- Nursing Care Plan: Subjective Data: Short Term IndependentDocument5 pagesNursing Care Plan: Subjective Data: Short Term IndependentIrish May SignioNo ratings yet
- Cognitive Behavioural Therapy For ADHD in Adults Systematic ReviewDocument9 pagesCognitive Behavioural Therapy For ADHD in Adults Systematic ReviewDana Ben SimonNo ratings yet
- Vaccine IngredientsDocument4 pagesVaccine IngredientsJulie BinghamNo ratings yet
- Concept Map of DMDocument1 pageConcept Map of DMRobert Timothy YapNo ratings yet
- Specialty Doctor/Senior Clinical Fellow in NEONATOLOGY (Middle Grade, ST4 To 8 Equivalent)Document10 pagesSpecialty Doctor/Senior Clinical Fellow in NEONATOLOGY (Middle Grade, ST4 To 8 Equivalent)madimadi11No ratings yet
- Stable Pasca Resusitasi TransportDocument76 pagesStable Pasca Resusitasi TransportDian KusumaNo ratings yet
- FEMM Charting Brochure (Bi-Fold)Document2 pagesFEMM Charting Brochure (Bi-Fold)teachingtashoilsNo ratings yet
- Proposal Optima Secure 73589641200000Document6 pagesProposal Optima Secure 73589641200000Star JasmineNo ratings yet
- Daily Pain Chart: Name Day DateDocument2 pagesDaily Pain Chart: Name Day DateAdiAri RosiuNo ratings yet
- Newborn Hearing Screening TestDocument11 pagesNewborn Hearing Screening TestGerardeanne Reposar100% (2)
- Drug Calculations 10th Brown Test BankDocument20 pagesDrug Calculations 10th Brown Test BankSaifoqq100% (1)
- Lesson Plan ON Upper Respiratory Tract Infections: Presented by Saranya.K Lecturer IconDocument26 pagesLesson Plan ON Upper Respiratory Tract Infections: Presented by Saranya.K Lecturer IconKumar BijayNo ratings yet
- After Activity Report Ems TrainingDocument9 pagesAfter Activity Report Ems TrainingMdrrmoPontevedranegoccNo ratings yet
- Presentation+Derma Needling+Power+Point+ 3Document20 pagesPresentation+Derma Needling+Power+Point+ 3Meireza Ajeng PratiwiNo ratings yet
- Harrisons Endocrinology 4Th Edition J Larry Jameson Full ChapterDocument57 pagesHarrisons Endocrinology 4Th Edition J Larry Jameson Full Chaptersally.kimberlin556100% (3)
- Cancer-Surviving Against The Odds - Brian CoxDocument58 pagesCancer-Surviving Against The Odds - Brian CoxwobblegobbleNo ratings yet
- Coronary Artery BypassDocument2 pagesCoronary Artery Bypassapi-341177338No ratings yet
- Basics of FundusDocument30 pagesBasics of FundusSamsam Almarez BacaltosNo ratings yet
- Set ADocument25 pagesSet AImraan KhanNo ratings yet
- JAAOS - Volume 11 - Issue 01 January & February 2003Document77 pagesJAAOS - Volume 11 - Issue 01 January & February 2003kenthepaNo ratings yet
- Medical Surgical Nursing Guillain Barre SyndromeDocument6 pagesMedical Surgical Nursing Guillain Barre SyndromeNoelyn BaluyanNo ratings yet
- MNG - D1 - 4. Rehabilitation Guideline of Myocardial Infarction PDFDocument83 pagesMNG - D1 - 4. Rehabilitation Guideline of Myocardial Infarction PDFRaagulan JeyashankerNo ratings yet
- Nurses Notes Date Focus Data Action Response: SubjectiveDocument2 pagesNurses Notes Date Focus Data Action Response: SubjectiveGILIANNE MARIE JIMENEANo ratings yet
- Group 4 - STKA BDocument4 pagesGroup 4 - STKA BNadia Izzatul AzkaNo ratings yet
- Campbell Incontinencia UrinariaDocument14 pagesCampbell Incontinencia UrinariaEmanuel FiggNo ratings yet