Download as pdf or txt
You might also like
- Clil Lesson Plan TemplateDocument5 pagesClil Lesson Plan Templatesandovalz24433% (3)
- The Gebusi - Chapter 3Document14 pagesThe Gebusi - Chapter 3Bob Jiggins67% (3)
- Technical Standard and Commentaries For Port and Harbour Facilities in JapanDocument66 pagesTechnical Standard and Commentaries For Port and Harbour Facilities in Japanrajmagi100% (4)
- Configuration 1 TDJ-709015DEI-90FDocument1 pageConfiguration 1 TDJ-709015DEI-90FДмитрий100% (1)
- Lydia Amir - Plato's Theory of LoveDocument9 pagesLydia Amir - Plato's Theory of Lovediegochillo5506No ratings yet
- Indian Plastic Industry, Plastic Industry in India, Plastic Industry, Plastic IndustriesDocument5 pagesIndian Plastic Industry, Plastic Industry in India, Plastic Industry, Plastic IndustriesPriyanshu ChokhaniNo ratings yet
- Light Is A Positive Regulator of Strigolactone Levels in Tomato RootsDocument4 pagesLight Is A Positive Regulator of Strigolactone Levels in Tomato RootslucasgirioNo ratings yet
- An Ancestral Role For CONSTITUTIVE TRIPLE RESPONSE1 Proteins in Both Ethylene and Abscisic Acid SignalingDocument16 pagesAn Ancestral Role For CONSTITUTIVE TRIPLE RESPONSE1 Proteins in Both Ethylene and Abscisic Acid SignalingMariz SintahaNo ratings yet
- 10 1093@aob@mcz100Document62 pages10 1093@aob@mcz100Dashiel ChevalierNo ratings yet
- Guard Cell Photosynthesis Is Critical For Stomatal Turgor Production, Yet Does Not Directly Mediate CO2 - and ABA-induced Stomatal ClosingDocument15 pagesGuard Cell Photosynthesis Is Critical For Stomatal Turgor Production, Yet Does Not Directly Mediate CO2 - and ABA-induced Stomatal Closingonefish1129No ratings yet
- Pala 2019Document9 pagesPala 2019rocio fernandez san juanNo ratings yet
- Chen 2012-Enraizamento de ArrosDocument7 pagesChen 2012-Enraizamento de ArrosMarcelo SonegoNo ratings yet
- Isolation and Antioxidant Activity of Flavonoids From HolarrhenaDocument6 pagesIsolation and Antioxidant Activity of Flavonoids From HolarrhenaIra MonicaNo ratings yet
- Jurnal AuksinDocument13 pagesJurnal AuksinKhina ChyaNkk KhirahNo ratings yet
- Rodriguez Pez Cebra 3BP-2015Document5 pagesRodriguez Pez Cebra 3BP-2015Gabriela RodriguezNo ratings yet
- Antioxidant and Hepatoprotective Effects of Different Eth - 2017 - South AfricanDocument7 pagesAntioxidant and Hepatoprotective Effects of Different Eth - 2017 - South AfricanGina LoveNo ratings yet
- Reproductive Development Modulates Gene Expression and Metabolite Levels With Possible Feedback Inhibition of Artemisinin in Artemisia AnnuaDocument11 pagesReproductive Development Modulates Gene Expression and Metabolite Levels With Possible Feedback Inhibition of Artemisinin in Artemisia AnnuanimaukNo ratings yet
- The TOPLESS Interactome A Framework For Gene RepreDocument17 pagesThe TOPLESS Interactome A Framework For Gene RepreBrun0No ratings yet
- Cytokinin Cross-Talking During Biotic and Abiotic Stress ResponsesDocument11 pagesCytokinin Cross-Talking During Biotic and Abiotic Stress ResponsesasifNo ratings yet
- JasmonatesDocument38 pagesJasmonatesSpinu LiliaNo ratings yet
- Structural and Functional ChangesDocument9 pagesStructural and Functional ChangesMAGALI GONZÁLEZ RODRÍGUEZNo ratings yet
- Chormova D., 2015x2Document17 pagesChormova D., 2015x2CarlosCifuentesNo ratings yet
- Pharmacological Activities of Plectranthus Cyclooxygenase and Xanthine Oxidase EnzymesDocument9 pagesPharmacological Activities of Plectranthus Cyclooxygenase and Xanthine Oxidase EnzymesYumnakarno 31No ratings yet
- Auxin-Cytokinin Interaction Regulates Meristem Development: Ying-Hua Su, Yu-Bo Liu and Xian-Sheng ZhangDocument10 pagesAuxin-Cytokinin Interaction Regulates Meristem Development: Ying-Hua Su, Yu-Bo Liu and Xian-Sheng ZhangJose Alburqueque PastorNo ratings yet
- Lavy and Estelle 2016 Mechanisms of Auxin SignalingDocument4 pagesLavy and Estelle 2016 Mechanisms of Auxin SignalingJacqueline Capataz TafurNo ratings yet
- A Review of Occurrence of Pharmaceuticals in SedimDocument4 pagesA Review of Occurrence of Pharmaceuticals in SedimDarlianaMelloSouzaNo ratings yet
- Hydrogen Sulfide Inhibits Fruit Softening by Regulating Ethylene Synthesis and Signaling Pathway in Tomato (Solanum Lycopersicum)Document7 pagesHydrogen Sulfide Inhibits Fruit Softening by Regulating Ethylene Synthesis and Signaling Pathway in Tomato (Solanum Lycopersicum)Miguel Francisco Maticorena QuispeNo ratings yet
- Adverse Morphological Development in Embryonic Zebrafish Exposed To Environmental Concentrations of Contaminants Individually and in MixtureDocument13 pagesAdverse Morphological Development in Embryonic Zebrafish Exposed To Environmental Concentrations of Contaminants Individually and in MixtureTiago TorresNo ratings yet
- Esparrago ArticuloDocument10 pagesEsparrago ArticuloROXANA SARAVIA ORTIZNo ratings yet
- Developmental Biology: Ce Dric Finet, Yvon JaillaisDocument13 pagesDevelopmental Biology: Ce Dric Finet, Yvon Jaillaisalkhafi firdausNo ratings yet
- Auxin Production As An Integrator of Environmental Cues For Developmental Growth RegulationDocument12 pagesAuxin Production As An Integrator of Environmental Cues For Developmental Growth RegulationSylviaYanaNo ratings yet
- Arginina 2Document12 pagesArginina 2Daniela SepulvedaNo ratings yet
- GaulDocument11 pagesGaulEdwin Pasha Jr.No ratings yet
- This Content Downloaded From 39.41.147.101 On Thu, 27 May 2021 00:05:54 UTCDocument4 pagesThis Content Downloaded From 39.41.147.101 On Thu, 27 May 2021 00:05:54 UTCIrfan Ullah 116-FBAS/MSCHM/F20No ratings yet
- Food and Chemical Toxicology: Lijun Sun, Jianbao Zhang, Xiaoyun Lu, Liyu Zhang, Yali ZhangDocument8 pagesFood and Chemical Toxicology: Lijun Sun, Jianbao Zhang, Xiaoyun Lu, Liyu Zhang, Yali ZhangSri LakshmiNo ratings yet
- 2015 - Carlos Ollas - Jasmonoylisoleucineaccumulationisneededforabscisic (Retrieved 2015-11-22)Document14 pages2015 - Carlos Ollas - Jasmonoylisoleucineaccumulationisneededforabscisic (Retrieved 2015-11-22)esn_kNo ratings yet
- Postharvest Biology and Technology: ArticleinfoDocument7 pagesPostharvest Biology and Technology: ArticleinfoMarcos MotaNo ratings yet
- Agastache Rugosa Leaf Extract Inhibits The iNOS Expression in - CompressedDocument6 pagesAgastache Rugosa Leaf Extract Inhibits The iNOS Expression in - Compressedela.sofiaNo ratings yet
- J Dent Res 2011 Takahashi 1463 8Document6 pagesJ Dent Res 2011 Takahashi 1463 8abcder1234No ratings yet
- Indole-3-Acetic Acid in Plant - Microbe InteractionsDocument41 pagesIndole-3-Acetic Acid in Plant - Microbe InteractionsMark Anthony RamosNo ratings yet
- Pro LinaDocument14 pagesPro LinaDaniela SepulvedaNo ratings yet
- Environmental and Experimental Botany: Haidong Li, Yun Wang, Jing Xiao, Kun XuDocument7 pagesEnvironmental and Experimental Botany: Haidong Li, Yun Wang, Jing Xiao, Kun XuArbi Iman Bahtiar NobizNo ratings yet
- Ecological Influence On Selected Aloe Vera Populations in Two Geographical Zones in NigeriaDocument6 pagesEcological Influence On Selected Aloe Vera Populations in Two Geographical Zones in Nigeriaibrahimflora7404No ratings yet
- Molluscicidaleffectoftuba-Tuba (Jatrophacurcaslinn) Extracts On Golden Apple Snail (Pomaceacanaliculatalamarck)Document6 pagesMolluscicidaleffectoftuba-Tuba (Jatrophacurcaslinn) Extracts On Golden Apple Snail (Pomaceacanaliculatalamarck)DanaNo ratings yet
- The Sesquiterpene Lactone Polymatin B From Smallanthus SonchifoliusDocument8 pagesThe Sesquiterpene Lactone Polymatin B From Smallanthus SonchifoliusJose Alberto PbNo ratings yet
- Chen2017 - Morphological and Chemical Variation of Stemona Tuberosa From MEDICINAL USESDocument8 pagesChen2017 - Morphological and Chemical Variation of Stemona Tuberosa From MEDICINAL USESanascopelNo ratings yet
- HormonDocument9 pagesHormonImène Ben Salah SoltaniNo ratings yet
- Hayward 2009Document13 pagesHayward 2009Ema DonNo ratings yet
- Evolution and Structural Diversification of PILS Putative Auxin Carriers in Plantsputative Auxin Carriers in PlantsDocument13 pagesEvolution and Structural Diversification of PILS Putative Auxin Carriers in Plantsputative Auxin Carriers in PlantsMugurel I. FeraruNo ratings yet
- Searching For Allelopathic Effects of Submerged Macrophytes On Phytoplankton-State of The Art and Open QuestionsDocument12 pagesSearching For Allelopathic Effects of Submerged Macrophytes On Phytoplankton-State of The Art and Open QuestionsDaniel De FreitasNo ratings yet
- Anthocyanin Interactions DNADocument20 pagesAnthocyanin Interactions DNAzainal ahzabNo ratings yet
- Food Chemistry: Sang Min Kim, Suk Woo Kang, Je-Seung Jeon, Yu-Jin Jung, Woo-Ri Kim, Chul Young Kim, Byung-Hun UmDocument8 pagesFood Chemistry: Sang Min Kim, Suk Woo Kang, Je-Seung Jeon, Yu-Jin Jung, Woo-Ri Kim, Chul Young Kim, Byung-Hun UmHazel Kaye OcaNo ratings yet
- Hypoxia-Inducible Transcription Factors in Fish ExDocument14 pagesHypoxia-Inducible Transcription Factors in Fish Ex454e75376rfufjcbNo ratings yet
- 1 s2.0 S0144861712007485 MainDocument9 pages1 s2.0 S0144861712007485 MainconcursolwcNo ratings yet
- Article1380714726 - Marek Et AlDocument8 pagesArticle1380714726 - Marek Et AlannisapritasNo ratings yet
- 2010 Okamoto DISDocument9 pages2010 Okamoto DISSalwa NabilahNo ratings yet
- Thalia 3026 17863 1 PBDocument19 pagesThalia 3026 17863 1 PBSalomón Páramo DelgadilloNo ratings yet
- New Phytologist - 2013 - Shahzad - Plant Defensin Type 1 PDF1 Protein Promiscuity and Expression Variation Within TheDocument14 pagesNew Phytologist - 2013 - Shahzad - Plant Defensin Type 1 PDF1 Protein Promiscuity and Expression Variation Within Thedora guzmanNo ratings yet
- Optimizing Ba and Iba Concentrations For Micro Propagation of Spineless Yucca Yucca Elephantipes IJERTV8IS010004Document5 pagesOptimizing Ba and Iba Concentrations For Micro Propagation of Spineless Yucca Yucca Elephantipes IJERTV8IS010004SHUBHAM JIBHAKATENo ratings yet
- Protein/gzr 048Document12 pagesProtein/gzr 048Arshia NazirNo ratings yet
- Aquatic ToxicologyDocument10 pagesAquatic ToxicologyJuan Pablo Monsalve QuirozNo ratings yet
- 1 s2.0 S0176161712003963 MainDocument9 pages1 s2.0 S0176161712003963 MainQuenguan FranklinNo ratings yet
- Fluorosis Induces Endoplasmic Reticulum Stress and Apoptosis in Osteoblasts in VivoDocument8 pagesFluorosis Induces Endoplasmic Reticulum Stress and Apoptosis in Osteoblasts in VivoKenneth MNo ratings yet
- Branching Hormone Is Busy Both Underground and OvergroundDocument4 pagesBranching Hormone Is Busy Both Underground and OvergroundRoshni NairNo ratings yet
- Phytochemistry: Yanhui Meng, Amanda J. Krzysiak, Michael J. Durako, Jennifer I. Kunzelman, Jeffrey L.C. WrightDocument6 pagesPhytochemistry: Yanhui Meng, Amanda J. Krzysiak, Michael J. Durako, Jennifer I. Kunzelman, Jeffrey L.C. WrightPutri Zahra ArdiyanitaNo ratings yet
- Influence of FOX genes on aging and aging-associated diseasesFrom EverandInfluence of FOX genes on aging and aging-associated diseasesNo ratings yet
- Global Change Biology - 2022 - Deines - Recent Cover Crop Adoption Is Associated With Small Maize and Soybean Yield LossesDocument14 pagesGlobal Change Biology - 2022 - Deines - Recent Cover Crop Adoption Is Associated With Small Maize and Soybean Yield LosseslucasgirioNo ratings yet
- Enhancing Citrus Fruit Yield Investigations Through Flight Height Optimization With UAV ImagingDocument9 pagesEnhancing Citrus Fruit Yield Investigations Through Flight Height Optimization With UAV ImaginglucasgirioNo ratings yet
- Remotesensing 16 00290 v2Document18 pagesRemotesensing 16 00290 v2lucasgirioNo ratings yet
- JIPB - 2022 - Wang - GIGANTEA Orthologs E2 Members Redundantly Determine Photoperiodic Flowering and Yield in SoybeanDocument15 pagesJIPB - 2022 - Wang - GIGANTEA Orthologs E2 Members Redundantly Determine Photoperiodic Flowering and Yield in SoybeanlucasgirioNo ratings yet
- Land 12 00584 v3Document26 pagesLand 12 00584 v3lucasgirioNo ratings yet
- 3Document292 pages3lucasgirioNo ratings yet
- Agriculture 13 00110Document21 pagesAgriculture 13 00110lucasgirioNo ratings yet
- Crop Science - 2023 - Egli - Yield Improvement and Yield Components A Comparison of Corn and SoybeanDocument11 pagesCrop Science - 2023 - Egli - Yield Improvement and Yield Components A Comparison of Corn and SoybeanlucasgirioNo ratings yet
- Global Change Biology - 2022 - Nendel - Future Area Expansion Outweighs Increasing Drought Risk For Soybean in EuropeDocument19 pagesGlobal Change Biology - 2022 - Nendel - Future Area Expansion Outweighs Increasing Drought Risk For Soybean in EuropelucasgirioNo ratings yet
- Plant Cell Environment - 2022 - Pelech - Leaf Plant To Canopy A Mechanistic Study On Aboveground Plasticity and PlantDocument17 pagesPlant Cell Environment - 2022 - Pelech - Leaf Plant To Canopy A Mechanistic Study On Aboveground Plasticity and PlantlucasgirioNo ratings yet
- Jac92222 232Document12 pagesJac92222 232lucasgirioNo ratings yet
- Application of Micronutrients As P-Fertilizer Coatings For Precision Distribution Improves Nutrients Use Efficiency in Soybean Crop EnvironmentDocument18 pagesApplication of Micronutrients As P-Fertilizer Coatings For Precision Distribution Improves Nutrients Use Efficiency in Soybean Crop EnvironmentlucasgirioNo ratings yet
- Agronomy 13 00170 v2Document14 pagesAgronomy 13 00170 v2lucasgirioNo ratings yet
- Fermentation 09 00141 v2Document13 pagesFermentation 09 00141 v2lucasgirioNo ratings yet
- 1 s2.0 S277237552200065X MainDocument9 pages1 s2.0 S277237552200065X MainlucasgirioNo ratings yet
- Advanced Science - 2021 - Zhou - Oat Plant Amyloids For Sustainable Functional MaterialsDocument11 pagesAdvanced Science - 2021 - Zhou - Oat Plant Amyloids For Sustainable Functional MaterialslucasgirioNo ratings yet
- Agriculture 12 00908Document15 pagesAgriculture 12 00908lucasgirioNo ratings yet
- 2022Document18 pages2022lucasgirioNo ratings yet
- Signalling Strategies During Drought and Salinity, Recent NewsDocument25 pagesSignalling Strategies During Drought and Salinity, Recent NewslucasgirioNo ratings yet
- 1 PBDocument12 pages1 PBlucasgirioNo ratings yet
- Initial Growth of Schizolobium Parahybae in Brazilian Cerrado Soil Under Liming and Mineral FertilizationDocument5 pagesInitial Growth of Schizolobium Parahybae in Brazilian Cerrado Soil Under Liming and Mineral FertilizationlucasgirioNo ratings yet
- Ion Transport in Halophytes: School of Agricultural Science, University of Tasmania, Hobart, Tas, AustraliaDocument49 pagesIon Transport in Halophytes: School of Agricultural Science, University of Tasmania, Hobart, Tas, AustralialucasgirioNo ratings yet
- TEXTO 3. Pesquisa Aplicada (Dupla)Document7 pagesTEXTO 3. Pesquisa Aplicada (Dupla)lucasgirioNo ratings yet
- Macronutrients in Watermelon Plants Fertilized With Potassium and Cattle ManureDocument5 pagesMacronutrients in Watermelon Plants Fertilized With Potassium and Cattle ManurelucasgirioNo ratings yet
- Nutrition and Yield of Gigante' Cactus Pear Cultivated With Different Spacings and Organic FertilizerDocument6 pagesNutrition and Yield of Gigante' Cactus Pear Cultivated With Different Spacings and Organic FertilizerlucasgirioNo ratings yet
- TEXTO 2. Pesquisa Aplicada (Dupla)Document5 pagesTEXTO 2. Pesquisa Aplicada (Dupla)lucasgirioNo ratings yet
- Yield of Sugarcane Submitted To Nitrogen Fertilization and Water Depths by Subsurface Drip IrrigationDocument6 pagesYield of Sugarcane Submitted To Nitrogen Fertilization and Water Depths by Subsurface Drip IrrigationlucasgirioNo ratings yet
- TEXTO 1. Estudo de Caso (Dupla)Document5 pagesTEXTO 1. Estudo de Caso (Dupla)lucasgirioNo ratings yet
- Doses and Application Methods of Azospirillum Brasilense in Irrigated Upland RiceDocument6 pagesDoses and Application Methods of Azospirillum Brasilense in Irrigated Upland RicelucasgirioNo ratings yet
- Growth and Mineral Nutrition of Baru (Dipteryx Alata Vogel) in Nutrient SolutionDocument6 pagesGrowth and Mineral Nutrition of Baru (Dipteryx Alata Vogel) in Nutrient SolutionlucasgirioNo ratings yet
- Intro To Signal Processing 2021Document510 pagesIntro To Signal Processing 2021crypto fanbaby0% (1)
- Organic Impurities in Fine Aggregates For Concrete: Standard Test Method ForDocument3 pagesOrganic Impurities in Fine Aggregates For Concrete: Standard Test Method ForKlisman A. Flores DurandNo ratings yet
- Cartridge English PDFDocument20 pagesCartridge English PDFkunto prNo ratings yet
- 02 - 01 Load Roadmap - FMS NG and Adv Features - Rev1Document62 pages02 - 01 Load Roadmap - FMS NG and Adv Features - Rev1Victor Borioli YancovitzNo ratings yet
- Study Guide: Kawabata Yasunari "Pomegranate" (Zakuro)Document8 pagesStudy Guide: Kawabata Yasunari "Pomegranate" (Zakuro)Beholdmyswarthyface75% (4)
- EME PPT Unit-5 - ElectiveDocument37 pagesEME PPT Unit-5 - ElectivesudeepkoreaNo ratings yet
- Role Responsibilities of Radiographer NewDocument49 pagesRole Responsibilities of Radiographer Newjiminsarah88No ratings yet
- Composites: Part BDocument13 pagesComposites: Part BShahnewaz BhuiyanNo ratings yet
- Phytosome: Presented byDocument14 pagesPhytosome: Presented bySari RamadhaniNo ratings yet
- Tutorial 4Document4 pagesTutorial 4Phoebe FaithNo ratings yet
- Wobenzym PSDocument3 pagesWobenzym PSManish PrakashNo ratings yet
- Acknowledgement: ReferencesDocument7 pagesAcknowledgement: Referencesdwayne420No ratings yet
- GeneralPhysics1 - Q2 - Mod2Lesson1Newton's Law of Universal Gravitation, Gravitational Field, and Gravitational Potential EnergyDocument28 pagesGeneralPhysics1 - Q2 - Mod2Lesson1Newton's Law of Universal Gravitation, Gravitational Field, and Gravitational Potential EnergyJose GulitiwNo ratings yet
- Iso1 Serie 581 PDFDocument231 pagesIso1 Serie 581 PDFchuylemalo1990No ratings yet
- THE BANAO BODONG ASSOCIATION (BBA) SMALL SCALE GOLD MINING IN GA-ANG MINES TALALANG, BALBALAN KALINGA CAR, PHILIPPINES by ROYCE LINGBAWANDocument22 pagesTHE BANAO BODONG ASSOCIATION (BBA) SMALL SCALE GOLD MINING IN GA-ANG MINES TALALANG, BALBALAN KALINGA CAR, PHILIPPINES by ROYCE LINGBAWANpopsky100% (1)
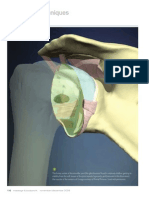
- The Rotator Cuff (Myofascial Techniques)Document4 pagesThe Rotator Cuff (Myofascial Techniques)Advanced-Trainings.com100% (3)
- PFCR ManualDocument777 pagesPFCR ManualFlo WheelerNo ratings yet
- Critical Functions & Design Criteria For Geogrids Used in Reinforced Aggregate Raft Foundations For Christchurch Rebuilt - Jan 2013Document4 pagesCritical Functions & Design Criteria For Geogrids Used in Reinforced Aggregate Raft Foundations For Christchurch Rebuilt - Jan 2013Victor WongNo ratings yet
- Pidari Lounge Menu 2021Document21 pagesPidari Lounge Menu 2021Akta SektiawanNo ratings yet
- Corporate Presentation Oramed December 2022Document41 pagesCorporate Presentation Oramed December 2022Самсон КукушкинNo ratings yet
- Mirroring Men and Masculinity in Joanne Harris' Chocolat and The Lollipop ShoesDocument50 pagesMirroring Men and Masculinity in Joanne Harris' Chocolat and The Lollipop ShoesBasneen Hudha AshrafNo ratings yet
- Final kw-4Document59 pagesFinal kw-4Chethan GowdaNo ratings yet
- Full Download Test Bank For Management 14th Edition John R Schermerhorn JR Daniel G Bachrach PDF Full ChapterDocument36 pagesFull Download Test Bank For Management 14th Edition John R Schermerhorn JR Daniel G Bachrach PDF Full Chaptercrincumose.at2d100% (26)
- Mikyas. 2020. Positive Science or Interpretive UnderstandingDocument16 pagesMikyas. 2020. Positive Science or Interpretive UnderstandingMikyas AberaNo ratings yet