
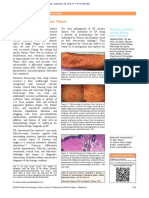
Sebaceous Gland Hair Shaft and Epidermal Barrier A
Sebaceous Gland Hair Shaft and Epidermal Barrier A
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5824)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Buku Ajar Psikiatri FKUI Ed 3Document638 pagesBuku Ajar Psikiatri FKUI Ed 3daily of sinta fu80% (20)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Buku Ajar Psikiatri FKUI Ed 3Document638 pagesBuku Ajar Psikiatri FKUI Ed 3daily of sinta fu0% (1)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- 553 1698 1 SMDocument7 pages553 1698 1 SMdaily of sinta fuNo ratings yet
- 299-Article Text-633-1-10-20200912Document5 pages299-Article Text-633-1-10-20200912daily of sinta fuNo ratings yet
- Contri Butions of The Psychosoc I A L SC Iences: 2.1 Jean Piaget and Cognitive DevelopmentDocument38 pagesContri Butions of The Psychosoc I A L SC Iences: 2.1 Jean Piaget and Cognitive Developmentdaily of sinta fuNo ratings yet
- C L Ass Ification Psych Iatry: Relation To Icd-1 0Document11 pagesC L Ass Ification Psych Iatry: Relation To Icd-1 0daily of sinta fuNo ratings yet
- International Journal of Anesthetics and Anesthesiology Ijaa 6 095Document5 pagesInternational Journal of Anesthetics and Anesthesiology Ijaa 6 095daily of sinta fuNo ratings yet
- International Journal of Anesthetics and Anesthesiology Ijaa 6 096Document5 pagesInternational Journal of Anesthetics and Anesthesiology Ijaa 6 096daily of sinta fuNo ratings yet
- Anesthetics and AnesthesiologyDocument3 pagesAnesthetics and Anesthesiologydaily of sinta fuNo ratings yet
- Dermoscopy of Keratosis PilarisDocument2 pagesDermoscopy of Keratosis Pilarisdaily of sinta fuNo ratings yet
- Untitled DesignDocument20 pagesUntitled Designdaily of sinta fuNo ratings yet
- Wang-Orlow2018 Article KeratosisPilarisAndItsSubtypesDocument25 pagesWang-Orlow2018 Article KeratosisPilarisAndItsSubtypesdaily of sinta fuNo ratings yet
- Fractional Carbon Dioxide Laser For Keratosis PilaDocument8 pagesFractional Carbon Dioxide Laser For Keratosis Piladaily of sinta fuNo ratings yet
- Keratosis Pilaris Rubra and Keratosis PiDocument5 pagesKeratosis Pilaris Rubra and Keratosis Pidaily of sinta fuNo ratings yet
- Imatinib Induced Keratosis Pilaris in ADocument4 pagesImatinib Induced Keratosis Pilaris in Adaily of sinta fuNo ratings yet
- Microbiology Reviewer: HistoryDocument29 pagesMicrobiology Reviewer: HistoryJOHN ROCKFORD LAZARONo ratings yet
- MCQS For SRDocument10 pagesMCQS For SRVivek BhatNo ratings yet
- Exam2006v27 05Document116 pagesExam2006v27 05akashkumarpanwarNo ratings yet
- Bacteriology-Block1-Lecture QuestionsDocument11 pagesBacteriology-Block1-Lecture QuestionsToni-Krys HardyNo ratings yet
- Hiv/Aids: Mwangi KJ (MSC Human Anat)Document47 pagesHiv/Aids: Mwangi KJ (MSC Human Anat)Diana Moraah MoturiNo ratings yet
- MalariaDocument9 pagesMalariaNitesh Bhura100% (5)
- Dédiés A15/A25: Designations Code Cond Prix H.TDocument4 pagesDédiés A15/A25: Designations Code Cond Prix H.TrhodesNo ratings yet
- Domains & Kingdoms of LifeDocument14 pagesDomains & Kingdoms of LifeRam Iyer100% (1)
- Cadenas Epidemiologic AsDocument51 pagesCadenas Epidemiologic AsYuliana De La CruzNo ratings yet
- Special StainsDocument27 pagesSpecial Stainsambadepravin100% (1)
- Buhner - Herbal Antivirals For Cov-19Document19 pagesBuhner - Herbal Antivirals For Cov-19seektherunes24100% (5)
- Annex 3: Recommendations To Assure The Quality, Safety and Efficacy of BCG VaccinesDocument50 pagesAnnex 3: Recommendations To Assure The Quality, Safety and Efficacy of BCG VaccinesDemo 1002No ratings yet
- Methods of Classification and Identifying MicroorganismsDocument3 pagesMethods of Classification and Identifying MicroorganismsBecca TayNo ratings yet
- Diagnostic Value of Candida Mannan Antigen and Anti-Mannan Igg, Igm Antibodies For Candida InfectionDocument18 pagesDiagnostic Value of Candida Mannan Antigen and Anti-Mannan Igg, Igm Antibodies For Candida Infectionmagda SiraitNo ratings yet
- Mechanism of p53 Dependent ApoptosisDocument5 pagesMechanism of p53 Dependent Apoptosischristevydinga8No ratings yet
- DAFTAR PUSTAKA Serologi Dan ImunologiDocument2 pagesDAFTAR PUSTAKA Serologi Dan ImunologiSari Anggun NNo ratings yet
- Febrele HemoragiceDocument23 pagesFebrele HemoragiceDan SlivinschiiNo ratings yet
- VPA & Glia &TLR Deckmann-2018-Autism-ADocument15 pagesVPA & Glia &TLR Deckmann-2018-Autism-AAhmed NagehNo ratings yet
- Turkish Health Exam Basci-April 2016Document56 pagesTurkish Health Exam Basci-April 2016Ali Jama100% (2)
- Test Bank For Foundations in Microbiology 8th Edition Kathleen P TalaroDocument14 pagesTest Bank For Foundations in Microbiology 8th Edition Kathleen P Talaromisbodensquallyr5g5pNo ratings yet
- What Is Genetic Testing Results For LifeDocument24 pagesWhat Is Genetic Testing Results For LiferamumyselfNo ratings yet
- Fimmu 13 853008Document15 pagesFimmu 13 853008hilaire0902No ratings yet
- EnterobacteriaceaeDocument31 pagesEnterobacteriaceaeVivek ChaudharyNo ratings yet
- Guillain Barre Syndrome A ReviewDocument7 pagesGuillain Barre Syndrome A ReviewEditor IJTSRDNo ratings yet
- Docshare - Tips - High Yield Biochemistrypdf PDFDocument41 pagesDocshare - Tips - High Yield Biochemistrypdf PDFArianne Joy C. TamarayNo ratings yet
- Reading Comprehension TextDocument2 pagesReading Comprehension TextMelanie Valeria Gualotuña GancinoNo ratings yet
- Fish ImmunoglobulinsDocument9 pagesFish ImmunoglobulinsSally ElshalqamyNo ratings yet
- GEN3051 Lecture 1: Human Genes and Human Genetic DisordersDocument5 pagesGEN3051 Lecture 1: Human Genes and Human Genetic DisordersAlessander Leyendecker JuniorNo ratings yet
- Chloramphenicol From WikipediaDocument9 pagesChloramphenicol From WikipediaMuhammad Miftahul HudaNo ratings yet
- TumorsDocument50 pagesTumorsdrshailaNo ratings yet




















































































Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5824)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Buku Ajar Psikiatri FKUI Ed 3Document638 pagesBuku Ajar Psikiatri FKUI Ed 3daily of sinta fu80% (20)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Buku Ajar Psikiatri FKUI Ed 3Document638 pagesBuku Ajar Psikiatri FKUI Ed 3daily of sinta fu0% (1)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- 553 1698 1 SMDocument7 pages553 1698 1 SMdaily of sinta fuNo ratings yet
- 299-Article Text-633-1-10-20200912Document5 pages299-Article Text-633-1-10-20200912daily of sinta fuNo ratings yet
- Contri Butions of The Psychosoc I A L SC Iences: 2.1 Jean Piaget and Cognitive DevelopmentDocument38 pagesContri Butions of The Psychosoc I A L SC Iences: 2.1 Jean Piaget and Cognitive Developmentdaily of sinta fuNo ratings yet
- C L Ass Ification Psych Iatry: Relation To Icd-1 0Document11 pagesC L Ass Ification Psych Iatry: Relation To Icd-1 0daily of sinta fuNo ratings yet
- International Journal of Anesthetics and Anesthesiology Ijaa 6 095Document5 pagesInternational Journal of Anesthetics and Anesthesiology Ijaa 6 095daily of sinta fuNo ratings yet
- International Journal of Anesthetics and Anesthesiology Ijaa 6 096Document5 pagesInternational Journal of Anesthetics and Anesthesiology Ijaa 6 096daily of sinta fuNo ratings yet
- Anesthetics and AnesthesiologyDocument3 pagesAnesthetics and Anesthesiologydaily of sinta fuNo ratings yet
- Dermoscopy of Keratosis PilarisDocument2 pagesDermoscopy of Keratosis Pilarisdaily of sinta fuNo ratings yet
- Untitled DesignDocument20 pagesUntitled Designdaily of sinta fuNo ratings yet
- Wang-Orlow2018 Article KeratosisPilarisAndItsSubtypesDocument25 pagesWang-Orlow2018 Article KeratosisPilarisAndItsSubtypesdaily of sinta fuNo ratings yet
- Fractional Carbon Dioxide Laser For Keratosis PilaDocument8 pagesFractional Carbon Dioxide Laser For Keratosis Piladaily of sinta fuNo ratings yet
- Keratosis Pilaris Rubra and Keratosis PiDocument5 pagesKeratosis Pilaris Rubra and Keratosis Pidaily of sinta fuNo ratings yet
- Imatinib Induced Keratosis Pilaris in ADocument4 pagesImatinib Induced Keratosis Pilaris in Adaily of sinta fuNo ratings yet
- Microbiology Reviewer: HistoryDocument29 pagesMicrobiology Reviewer: HistoryJOHN ROCKFORD LAZARONo ratings yet
- MCQS For SRDocument10 pagesMCQS For SRVivek BhatNo ratings yet
- Exam2006v27 05Document116 pagesExam2006v27 05akashkumarpanwarNo ratings yet
- Bacteriology-Block1-Lecture QuestionsDocument11 pagesBacteriology-Block1-Lecture QuestionsToni-Krys HardyNo ratings yet
- Hiv/Aids: Mwangi KJ (MSC Human Anat)Document47 pagesHiv/Aids: Mwangi KJ (MSC Human Anat)Diana Moraah MoturiNo ratings yet
- MalariaDocument9 pagesMalariaNitesh Bhura100% (5)
- Dédiés A15/A25: Designations Code Cond Prix H.TDocument4 pagesDédiés A15/A25: Designations Code Cond Prix H.TrhodesNo ratings yet
- Domains & Kingdoms of LifeDocument14 pagesDomains & Kingdoms of LifeRam Iyer100% (1)
- Cadenas Epidemiologic AsDocument51 pagesCadenas Epidemiologic AsYuliana De La CruzNo ratings yet
- Special StainsDocument27 pagesSpecial Stainsambadepravin100% (1)
- Buhner - Herbal Antivirals For Cov-19Document19 pagesBuhner - Herbal Antivirals For Cov-19seektherunes24100% (5)
- Annex 3: Recommendations To Assure The Quality, Safety and Efficacy of BCG VaccinesDocument50 pagesAnnex 3: Recommendations To Assure The Quality, Safety and Efficacy of BCG VaccinesDemo 1002No ratings yet
- Methods of Classification and Identifying MicroorganismsDocument3 pagesMethods of Classification and Identifying MicroorganismsBecca TayNo ratings yet
- Diagnostic Value of Candida Mannan Antigen and Anti-Mannan Igg, Igm Antibodies For Candida InfectionDocument18 pagesDiagnostic Value of Candida Mannan Antigen and Anti-Mannan Igg, Igm Antibodies For Candida Infectionmagda SiraitNo ratings yet
- Mechanism of p53 Dependent ApoptosisDocument5 pagesMechanism of p53 Dependent Apoptosischristevydinga8No ratings yet
- DAFTAR PUSTAKA Serologi Dan ImunologiDocument2 pagesDAFTAR PUSTAKA Serologi Dan ImunologiSari Anggun NNo ratings yet
- Febrele HemoragiceDocument23 pagesFebrele HemoragiceDan SlivinschiiNo ratings yet
- VPA & Glia &TLR Deckmann-2018-Autism-ADocument15 pagesVPA & Glia &TLR Deckmann-2018-Autism-AAhmed NagehNo ratings yet
- Turkish Health Exam Basci-April 2016Document56 pagesTurkish Health Exam Basci-April 2016Ali Jama100% (2)
- Test Bank For Foundations in Microbiology 8th Edition Kathleen P TalaroDocument14 pagesTest Bank For Foundations in Microbiology 8th Edition Kathleen P Talaromisbodensquallyr5g5pNo ratings yet
- What Is Genetic Testing Results For LifeDocument24 pagesWhat Is Genetic Testing Results For LiferamumyselfNo ratings yet
- Fimmu 13 853008Document15 pagesFimmu 13 853008hilaire0902No ratings yet
- EnterobacteriaceaeDocument31 pagesEnterobacteriaceaeVivek ChaudharyNo ratings yet
- Guillain Barre Syndrome A ReviewDocument7 pagesGuillain Barre Syndrome A ReviewEditor IJTSRDNo ratings yet
- Docshare - Tips - High Yield Biochemistrypdf PDFDocument41 pagesDocshare - Tips - High Yield Biochemistrypdf PDFArianne Joy C. TamarayNo ratings yet
- Reading Comprehension TextDocument2 pagesReading Comprehension TextMelanie Valeria Gualotuña GancinoNo ratings yet
- Fish ImmunoglobulinsDocument9 pagesFish ImmunoglobulinsSally ElshalqamyNo ratings yet
- GEN3051 Lecture 1: Human Genes and Human Genetic DisordersDocument5 pagesGEN3051 Lecture 1: Human Genes and Human Genetic DisordersAlessander Leyendecker JuniorNo ratings yet
- Chloramphenicol From WikipediaDocument9 pagesChloramphenicol From WikipediaMuhammad Miftahul HudaNo ratings yet
- TumorsDocument50 pagesTumorsdrshailaNo ratings yet