Download as pdf or txt
You might also like
- Gram Positive Bacteria: Antibacterial DrugsDocument4 pagesGram Positive Bacteria: Antibacterial DrugsMOHAMAD HASSOUNANo ratings yet
- Mcqs in Biochemistry: Danylo Halytskyi Lviv National Medical UniversityDocument320 pagesMcqs in Biochemistry: Danylo Halytskyi Lviv National Medical UniversityKatarzyna Rosemary Czerwińska100% (1)
- Biochemistry EnzymesDocument88 pagesBiochemistry EnzymesLama QaimariNo ratings yet
- Learning Objectives Identify The 6 Classes ofDocument21 pagesLearning Objectives Identify The 6 Classes ofhahmed78No ratings yet
- Enzymes: 5e Chapter 1: p20-27 6e Chapter 1: p21-25Document64 pagesEnzymes: 5e Chapter 1: p20-27 6e Chapter 1: p21-25mehNo ratings yet
- Enzymes: Dr. S.Chakravarty MDDocument76 pagesEnzymes: Dr. S.Chakravarty MDRaad GaffazNo ratings yet
- Module IIIDocument20 pagesModule IIIVishal MishraNo ratings yet
- Chapter Outline: and V and VDocument5 pagesChapter Outline: and V and VMae RabariaNo ratings yet
- Enz FactorsDocument25 pagesEnz FactorsSparrowNo ratings yet
- 04 Enz-FactorsDocument25 pages04 Enz-FactorsDeepthiNo ratings yet
- BIOTECH REPORT UpdatedDocument37 pagesBIOTECH REPORT UpdatedMaggie VillacortaNo ratings yet
- MetabolismDocument27 pagesMetabolismAnum RasoolNo ratings yet
- Lecture 4b. Chemical Kinetics 2020Document23 pagesLecture 4b. Chemical Kinetics 2020Montassar DridiNo ratings yet
- Chapter 2: Diagnostic Enzymology: E+S Es E+PDocument87 pagesChapter 2: Diagnostic Enzymology: E+S Es E+PDereje KasuNo ratings yet
- Lecture HandoutsDocument6 pagesLecture HandoutsMohamidin MamalapatNo ratings yet
- Lecture 3 Notes - EnzymologyDocument9 pagesLecture 3 Notes - EnzymologyYi Gong100% (3)
- Lecture 5a CatalysisDocument22 pagesLecture 5a CatalysisSandeep ChawlaNo ratings yet
- Enzyme PropertiesDocument45 pagesEnzyme PropertiesSarah De Los Santos100% (1)
- Enzymes: Protein CatalystsDocument15 pagesEnzymes: Protein Catalystsgmurthy_1No ratings yet
- Enzyme CatalysisDocument65 pagesEnzyme Catalysissushant100% (1)
- Bi515 Chapter 6, Part 1: Enzymes: Overview 2010Document22 pagesBi515 Chapter 6, Part 1: Enzymes: Overview 2010MattNo ratings yet
- Module 2 Enzyme TransDocument11 pagesModule 2 Enzyme TransKathleen CedilloNo ratings yet
- Voet - Chapt - 12 Properties of EnzymesDocument102 pagesVoet - Chapt - 12 Properties of Enzymestelmo flowNo ratings yet
- Enzymes LectureDocument115 pagesEnzymes LectureJane Tai100% (1)
- Voet Chapt 11Document112 pagesVoet Chapt 11Sandhya KanetkarNo ratings yet
- The Organic Chemistry of Enzyme-Catalysed ReactionsDocument63 pagesThe Organic Chemistry of Enzyme-Catalysed ReactionskunwarskNo ratings yet
- Enzymes LectureDocument120 pagesEnzymes LectureHERSEY MIAYONo ratings yet
- Catalytic Principles and Reaction Mechanisms of EnzymesDocument50 pagesCatalytic Principles and Reaction Mechanisms of EnzymesBalew GetaNo ratings yet
- TRK1 2013 Chapt 3 (Part 1)Document17 pagesTRK1 2013 Chapt 3 (Part 1)Yoel Dwi Putra GultomNo ratings yet
- Equations in Enzyme KineticsDocument29 pagesEquations in Enzyme KineticsAndrew Sam100% (2)
- CH 7 Kinetics and RegulationDocument8 pagesCH 7 Kinetics and Regulationcococat015No ratings yet
- C H O + 6O (G) 6co + 6H O: - Under Standard Conditions of 25° C and One Atmosphere Pressure, TheDocument42 pagesC H O + 6O (G) 6co + 6H O: - Under Standard Conditions of 25° C and One Atmosphere Pressure, TheErsilia VerlinghieriNo ratings yet
- BP U6 EnzymesDocument68 pagesBP U6 EnzymesChristian Angelo AgbunagNo ratings yet
- Enzymes WorksheetDocument2 pagesEnzymes WorksheetNatalie Pemberton100% (1)
- Enzyme Kinetics BasicDocument14 pagesEnzyme Kinetics Basicnibraz307No ratings yet
- Bai Giang Chuong 06Document36 pagesBai Giang Chuong 06Minh TâmNo ratings yet
- Chapter 2: Diagnostic Enzymology: E+S Es E+PDocument72 pagesChapter 2: Diagnostic Enzymology: E+S Es E+PQananiisa Lammii Gammada100% (1)
- Atkinson1968 PDFDocument5 pagesAtkinson1968 PDFOleggValVilNo ratings yet
- Chapter 20 EDocument20 pagesChapter 20 EBerru VardarNo ratings yet
- Enzymatic Catalysis by Asmaa Abouzaid, TA at Zewail City, Egypt.Document43 pagesEnzymatic Catalysis by Asmaa Abouzaid, TA at Zewail City, Egypt.asmaa abouzeidNo ratings yet
- T: 5.6 R E P: Opic Eaction Nergy RofileDocument4 pagesT: 5.6 R E P: Opic Eaction Nergy RofileJannah ElmaghrabyNo ratings yet
- Medical Biochemistry: Enzymes: Mr. Genaro F. Alderite JR, MsermDocument58 pagesMedical Biochemistry: Enzymes: Mr. Genaro F. Alderite JR, MsermNash DeniegaNo ratings yet
- Ivatio: ACT NDocument18 pagesIvatio: ACT NNikhil DeshmukhNo ratings yet
- Lecture 17-Multisubstrate Enzyme RXN KineticsDocument13 pagesLecture 17-Multisubstrate Enzyme RXN KineticsAmogha G C 1SI19CH002No ratings yet
- 5.enzymes CompleteDocument60 pages5.enzymes Completeazyhuang77No ratings yet
- Bioenergetics, Enzymes, MetabolismDocument101 pagesBioenergetics, Enzymes, MetabolismRhendiel SanchezNo ratings yet
- Bioenergetics, Enzymes, Metabolism PDFDocument85 pagesBioenergetics, Enzymes, Metabolism PDFRhendiel SanchezNo ratings yet
- Chapter 6.7 Abbreviated Lecture NotesDocument20 pagesChapter 6.7 Abbreviated Lecture NotesZach MaxwellNo ratings yet
- Weeks 2 and 3 - EnzymesDocument30 pagesWeeks 2 and 3 - EnzymesAndrei FloreaNo ratings yet
- ENZYMES NOTES (Recovered)Document10 pagesENZYMES NOTES (Recovered)zipporahwaithera404No ratings yet
- Enzyme LabDocument47 pagesEnzyme Labdentcu70100% (1)
- Nonisothermal Heterogeneous Reaction in A Denaturable Immobilized Enzyme CatalystDocument5 pagesNonisothermal Heterogeneous Reaction in A Denaturable Immobilized Enzyme CatalystbagyabioNo ratings yet
- Bio EnergeticsDocument24 pagesBio EnergeticsNouman SiddiqueNo ratings yet
- Reactor EngineeringDocument19 pagesReactor EngineeringAlvine AyietaNo ratings yet
- Chemistry Notes For Class 12 Chapter 4 Chemical KineticsDocument11 pagesChemistry Notes For Class 12 Chapter 4 Chemical KineticsAyush singh PrinceNo ratings yet
- ENZYME KINETICS Transcribed SlidesDocument8 pagesENZYME KINETICS Transcribed Slidest4gjzhpfjcNo ratings yet
- EnzymesDocument80 pagesEnzymesThisha MohanNo ratings yet
- BS 3Document60 pagesBS 3himanshu_agraNo ratings yet
- CH6 Enzyme Kinetics Spring2024Document78 pagesCH6 Enzyme Kinetics Spring2024Farah Abu AlshaarNo ratings yet
- 2.enzyme Kinetics 2021Document133 pages2.enzyme Kinetics 2021Amrita SinghNo ratings yet
- Glycogen Found In-: The Goals of This Lesson o o o Bioenergetics and ThermodynamicsDocument24 pagesGlycogen Found In-: The Goals of This Lesson o o o Bioenergetics and ThermodynamicsRuhulAminNo ratings yet
- Parasitic Diseases of The LungDocument27 pagesParasitic Diseases of The Lungsultan khabeebNo ratings yet
- Respiratory Tract MicrobiologyDocument68 pagesRespiratory Tract Microbiologysultan khabeebNo ratings yet
- Fungal Lung DiseasesDocument28 pagesFungal Lung Diseasessultan khabeebNo ratings yet
- TI1 and TI2Document4 pagesTI1 and TI2sultan khabeebNo ratings yet
- Cardiovascular System HistologyDocument45 pagesCardiovascular System Histologysultan khabeebNo ratings yet
- Antigen Antibody Interactions: Serology Test: Serological Diagnostic TestsDocument44 pagesAntigen Antibody Interactions: Serology Test: Serological Diagnostic Testssultan khabeebNo ratings yet
- 2 Respiratory Medicine: Streptococcus Pneumoniae Klebsiella Pneumoniae Legionella Pneumophila Mycoplasma PneumoniaeDocument24 pages2 Respiratory Medicine: Streptococcus Pneumoniae Klebsiella Pneumoniae Legionella Pneumophila Mycoplasma Pneumoniaesultan khabeeb100% (1)
- Drugs For HyperlipidemiaDocument29 pagesDrugs For Hyperlipidemiasultan khabeebNo ratings yet
- Cardiovascular System Histo LabDocument53 pagesCardiovascular System Histo Labsultan khabeebNo ratings yet
- Microbiology of CVSDocument44 pagesMicrobiology of CVSsultan khabeebNo ratings yet
- Drugs For Congestive Heart FailureDocument46 pagesDrugs For Congestive Heart Failuresultan khabeeb100% (1)
- Antihypertensive DrugsDocument56 pagesAntihypertensive Drugssultan khabeebNo ratings yet
- Citric Acid Cycle - Pyruvate DehydrogenaseDocument33 pagesCitric Acid Cycle - Pyruvate Dehydrogenasesultan khabeeb100% (1)
- Anti-Arrhythmic DrugsDocument45 pagesAnti-Arrhythmic Drugssultan khabeebNo ratings yet
- Antianginal DrugsDocument32 pagesAntianginal Drugssultan khabeebNo ratings yet
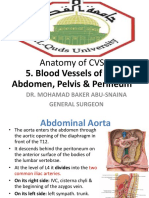
- Anatomy of CVS: 5. Blood Vessels of The Abdomen, Pelvis & PerineumDocument30 pagesAnatomy of CVS: 5. Blood Vessels of The Abdomen, Pelvis & Perineumsultan khabeebNo ratings yet
- CH 1 - Cell Injury III Subcellular Responses To Injury - PPT (Compatibility Mode)Document7 pagesCH 1 - Cell Injury III Subcellular Responses To Injury - PPT (Compatibility Mode)sultan khabeebNo ratings yet
- CH 1 - Cell Injury IV Reversible Vs Irreversible (Compatibility Mode)Document12 pagesCH 1 - Cell Injury IV Reversible Vs Irreversible (Compatibility Mode)sultan khabeebNo ratings yet
- The Citric Acid CycleDocument32 pagesThe Citric Acid Cyclesultan khabeebNo ratings yet
- Anatomy of CVS: 4. Blood Vessels of The Head & NeckDocument37 pagesAnatomy of CVS: 4. Blood Vessels of The Head & Necksultan khabeebNo ratings yet
- Anatomy of CVS: 2. The HeartDocument68 pagesAnatomy of CVS: 2. The Heartsultan khabeebNo ratings yet
- Anatomy of CVS: 3. Blood Vessels of The ThoraxDocument20 pagesAnatomy of CVS: 3. Blood Vessels of The Thoraxsultan khabeebNo ratings yet
- 1.1 Enzymology (Bravo)Document11 pages1.1 Enzymology (Bravo)Arman Carl DulayNo ratings yet
- Mending A Broken HeartDocument3 pagesMending A Broken Heartkatrina_velasco_1No ratings yet
- 2007, Volume 67, Issue 1Document152 pages2007, Volume 67, Issue 1Kareem JakouchNo ratings yet
- Cooling Tower REPORTDocument25 pagesCooling Tower REPORTSaroj Kumar100% (1)
- MetabolismDocument27 pagesMetabolismAnum RasoolNo ratings yet
- Corrosion Inhibition Ability of Shorea Robusta (Sakhu) Leaves)Document4 pagesCorrosion Inhibition Ability of Shorea Robusta (Sakhu) Leaves)Dr. Syed Khalid HasanNo ratings yet
- Enzymatic Cell Wall Degradation of Chlorella Vulgaris and Other Microalgae For Biofuels Production 2012Document15 pagesEnzymatic Cell Wall Degradation of Chlorella Vulgaris and Other Microalgae For Biofuels Production 2012Ryan Asyhari100% (1)
- Toxicology Course Notes 2009Document342 pagesToxicology Course Notes 2009paulheroux100% (5)
- Trial Sem 1 2017 SMK THREE RIVERS Answers PDFDocument12 pagesTrial Sem 1 2017 SMK THREE RIVERS Answers PDFChee Beng YeapNo ratings yet
- Docking of Ach and BuchDocument25 pagesDocking of Ach and BuchRISHITA CHAUHANNo ratings yet
- Combined Scale Removal and Scale Inhibition TreatmentsDocument6 pagesCombined Scale Removal and Scale Inhibition TreatmentsJu Bbin100% (2)
- 1st Year Biochemistry Guidance BookletDocument67 pages1st Year Biochemistry Guidance Bookletusama HafeezNo ratings yet
- MCQS CH 3 Bio Part 1 1Document2 pagesMCQS CH 3 Bio Part 1 1All kinds of information on channelNo ratings yet
- Fischer Et Al-2008-Journal of Pineal ResearchDocument15 pagesFischer Et Al-2008-Journal of Pineal ResearchgeoaislaNo ratings yet
- Lecture BiocidesDocument40 pagesLecture BiocidesRuth AkomohNo ratings yet
- C 1 1 2025 Topic Test MsDocument4 pagesC 1 1 2025 Topic Test MsRawanMazen SharifNo ratings yet
- Indomethacin From Anti-Inflammatory To Anticancer AgentDocument26 pagesIndomethacin From Anti-Inflammatory To Anticancer AgentYaqeen Alhaqq F. GhaziNo ratings yet
- D-Manoza FullDocument8 pagesD-Manoza FullSeulean BogdanNo ratings yet
- 6.3 Enzyme Kinetics LabDocument3 pages6.3 Enzyme Kinetics LabSiegNo ratings yet
- Inhibition of Anaerobic Digestion Process A ReviewDocument26 pagesInhibition of Anaerobic Digestion Process A ReviewCintia Andrade MoóNo ratings yet
- EnzymesDocument5 pagesEnzymesLea IoaNo ratings yet
- Biochemistry and Pharmacology of Reversible Inhibitors of MAO-Agents A Focus On MoclobemideDocument12 pagesBiochemistry and Pharmacology of Reversible Inhibitors of MAO-Agents A Focus On MoclobemideYoselin Milagros Chambi VelasquezNo ratings yet
- Ammonia ChannelingDocument11 pagesAmmonia ChannelingarivasudevaNo ratings yet
- Muegge 2010Document46 pagesMuegge 2010Santi SuronoNo ratings yet
- Toxicodynamics: and Molecular LevelsDocument24 pagesToxicodynamics: and Molecular LevelsRajabMumbeeNo ratings yet
- Objective Weed Science - Second EditionDocument40 pagesObjective Weed Science - Second EditionSurinder Singh RanaNo ratings yet
- Chapter 6 - The Behavior of Proteins ENZYMES-2018Document111 pagesChapter 6 - The Behavior of Proteins ENZYMES-2018ruaa mhmadNo ratings yet
- Carbohydrate Based Molecular ScaffoldingDocument43 pagesCarbohydrate Based Molecular ScaffoldingAngelo GNo ratings yet