Download as pdf or txt
You might also like
- CTS Case Study Talc JJDocument5 pagesCTS Case Study Talc JJ2 secNo ratings yet
- NF-JB and Cancer: How Intimate Is This Relationship: Ó Springer Science+Business Media, LLC. 2009Document13 pagesNF-JB and Cancer: How Intimate Is This Relationship: Ó Springer Science+Business Media, LLC. 2009rendyjiwonoNo ratings yet
- Ijms 22 08223Document19 pagesIjms 22 08223Giovanni Aleksey GuersonNo ratings yet
- Role of EbvDocument6 pagesRole of EbvMuhamad Ansori BastianNo ratings yet
- NF KB ThesisDocument6 pagesNF KB Thesisjessicaspellfayetteville100% (2)
- Plos One Lmp1Document23 pagesPlos One Lmp1Karol BalsecaNo ratings yet
- 2022-oncogene-UPF1:circRPPH1:ATF3 Feedback Loop Promotes The Malignant Phenotype and Stemness of GSCsDocument15 pages2022-oncogene-UPF1:circRPPH1:ATF3 Feedback Loop Promotes The Malignant Phenotype and Stemness of GSCszhe zhNo ratings yet
- Restoration of p53 Expression in Human Cancer CellDocument9 pagesRestoration of p53 Expression in Human Cancer CellnsainagachaitanyaNo ratings yet
- Clin Cancer Res 2009 15 425.fullDocument6 pagesClin Cancer Res 2009 15 425.fullthanosssNo ratings yet
- Nedd4-Binding Protein 1 and TNFAIP3-Interacting Protein 1 Control MHC-1 Display in NeuroblastomaDocument11 pagesNedd4-Binding Protein 1 and TNFAIP3-Interacting Protein 1 Control MHC-1 Display in NeuroblastomalauraNo ratings yet
- HHS Public AccessDocument17 pagesHHS Public AccessSanthiya KunasegaranNo ratings yet
- Nuclear Trafficking in Health and Disease: SciencedirectDocument8 pagesNuclear Trafficking in Health and Disease: SciencedirectMariaNo ratings yet
- Nf-Κb-Modulated Mir-130A Targets Tnf-Α In Cervical Cancer CellsDocument14 pagesNf-Κb-Modulated Mir-130A Targets Tnf-Α In Cervical Cancer CellsMazhar HussainNo ratings yet
- 2019-Sanghv-Glycolation NRF2Document35 pages2019-Sanghv-Glycolation NRF2HaiNo ratings yet
- Bagus BangetDocument12 pagesBagus BangetSyiefa RenandaNo ratings yet
- Dogma CentralDocument20 pagesDogma Centraldwinov educationNo ratings yet
- Gastroenterohepatology 1Document33 pagesGastroenterohepatology 1Laboratorium Ansari SalehNo ratings yet
- Neuropilins Intumor Biology: Molecular PathwaysDocument5 pagesNeuropilins Intumor Biology: Molecular PathwaysdupuytrenNo ratings yet
- RIP Kinases: Key Decision Makers in Cell Death and Innate ImmunityDocument12 pagesRIP Kinases: Key Decision Makers in Cell Death and Innate ImmunityBalarama Krishna GogineniNo ratings yet
- Brain-Derived Neurotrophic Factor in Gastroenterology Oncology - Short Review of Current LiteratureDocument5 pagesBrain-Derived Neurotrophic Factor in Gastroenterology Oncology - Short Review of Current LiteratureAhana MukherjeeNo ratings yet
- 2010NFkB抑制剂大总结Identification of known drugs that act as inhibitors of NF-kappaB signaling and their mechanism of action.Document23 pages2010NFkB抑制剂大总结Identification of known drugs that act as inhibitors of NF-kappaB signaling and their mechanism of action.meltwithsnow163.comNo ratings yet
- The Retinoblastoma Tumor-Suppressor Gene, The Exception That Proves The RuleDocument11 pagesThe Retinoblastoma Tumor-Suppressor Gene, The Exception That Proves The RuleSanthiya KunasegaranNo ratings yet
- Presentación 27Document18 pagesPresentación 27Felipe RayoNo ratings yet
- Tumor Suppressor Genes: PatofisiologiDocument17 pagesTumor Suppressor Genes: Patofisiologidr_demaNo ratings yet
- Castro, 2022, Targeting The Complex Protein Network of MYCN-amplified Anaplastic EpendymomaDocument14 pagesCastro, 2022, Targeting The Complex Protein Network of MYCN-amplified Anaplastic EpendymomadhanNo ratings yet
- Conf Munique 2012 - Professor Serge JurasunasDocument88 pagesConf Munique 2012 - Professor Serge Jurasunasnomehoda513No ratings yet
- 6354 FullDocument11 pages6354 FullHector Javier BurgosNo ratings yet
- Unravel The Molecular Mechanism of XBP1 in Regulating The Biology of Cancer CellsDocument12 pagesUnravel The Molecular Mechanism of XBP1 in Regulating The Biology of Cancer CellsDavi MuhammadNo ratings yet
- The Association and Signi Cance of p53 in Gynecologic Cancers: The Potential of Targeted TherapyDocument16 pagesThe Association and Signi Cance of p53 in Gynecologic Cancers: The Potential of Targeted TherapychasingNo ratings yet
- Diagnostic Review of Neurofibromatosis Type 1: Pathology Case Reviews January 2014Document10 pagesDiagnostic Review of Neurofibromatosis Type 1: Pathology Case Reviews January 2014Xavier PeñaloNo ratings yet
- Alteration of Epigenetic Regulation by Long NoncodDocument16 pagesAlteration of Epigenetic Regulation by Long NoncodVY TRẦN ĐOÀN NHẬTNo ratings yet
- Upregulation of Mir-107 Expression Following Hyperbaric Oxygen Treatment Suppresses Hmgb1/Rage Signaling in Degenerated Human Nucleus Pulposus CellsDocument14 pagesUpregulation of Mir-107 Expression Following Hyperbaric Oxygen Treatment Suppresses Hmgb1/Rage Signaling in Degenerated Human Nucleus Pulposus CellsgeorgiaNo ratings yet
- TP53 Mutations Upregulate RCP Expression Via Sp1/3 To Drive Lung Cancer ProgressionDocument15 pagesTP53 Mutations Upregulate RCP Expression Via Sp1/3 To Drive Lung Cancer ProgressionValen EstevezNo ratings yet
- Jonathan M. Fahey, Jennifer S. Stancill, Brian C. Smith, and Albert W. GirottiDocument36 pagesJonathan M. Fahey, Jennifer S. Stancill, Brian C. Smith, and Albert W. GirottiRizki Dwi PutriNo ratings yet
- c-MYC-directed NRF2 Drives Malignant Progression of Head and Neck Cancer Via Glucose-6-Phosphate Dehydrogenase and Transketolase ActivationDocument16 pagesc-MYC-directed NRF2 Drives Malignant Progression of Head and Neck Cancer Via Glucose-6-Phosphate Dehydrogenase and Transketolase ActivationsirfengchenNo ratings yet
- BCH4125 2022 - Lecture6 Jan27Document27 pagesBCH4125 2022 - Lecture6 Jan27Fatima AkbariNo ratings yet
- To Be, or Not To Be: NF-JB Is The Answer - Role of Rel/NF-jB in The Regulation of ApoptosisDocument22 pagesTo Be, or Not To Be: NF-JB Is The Answer - Role of Rel/NF-jB in The Regulation of ApoptosisPrinzessinElaineNo ratings yet
- Abdul Rafay Javed Roll No 23Document9 pagesAbdul Rafay Javed Roll No 23Rafay MirzaNo ratings yet
- Halliday2019 Article EmergingTargetedTherapiesForThDocument12 pagesHalliday2019 Article EmergingTargetedTherapiesForThAlfiah NoorNo ratings yet
- 1 s2.0 S0145212612004456 MainDocument9 pages1 s2.0 S0145212612004456 MainMilana PerkovićNo ratings yet
- Art de OncoDocument15 pagesArt de OncoGabriela SchettiniNo ratings yet
- 2007 Article 63Document5 pages2007 Article 63marcussiNo ratings yet
- 2017 FBW7 Mutations Mediate ResistanceDocument10 pages2017 FBW7 Mutations Mediate ResistancemaomaochongNo ratings yet
- Porte Tal 18Document18 pagesPorte Tal 18alfredoibcNo ratings yet
- Tumor Necrosis Factor and Cancer, Buddies or Foes?Document14 pagesTumor Necrosis Factor and Cancer, Buddies or Foes?hoangphuong08101992No ratings yet
- Role of NF-Binthe Regulation of Immunity and Apoptosis: Kavitha BharathamDocument30 pagesRole of NF-Binthe Regulation of Immunity and Apoptosis: Kavitha BharathamArief Adi SaputroNo ratings yet
- P 53 PathwayDocument10 pagesP 53 PathwayShera NeazNo ratings yet
- Perineural Invasion of Cancer A Complex Crosstalk Between Cells and Molecules in The Perineural NicheDocument21 pagesPerineural Invasion of Cancer A Complex Crosstalk Between Cells and Molecules in The Perineural NichegangligonNo ratings yet
- 2018 - STAT3 in Regulatory T Cells Are PD-1 Blockade in Resected Metastatic MelanomaDocument12 pages2018 - STAT3 in Regulatory T Cells Are PD-1 Blockade in Resected Metastatic Melanoma1262615286No ratings yet
- Review Article: Recent Advances in p53 Research and Cancer TreatmentDocument8 pagesReview Article: Recent Advances in p53 Research and Cancer TreatmentSanthiya KunasegaranNo ratings yet
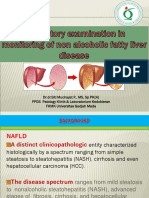
- Review Article: Signal Transductions and Nonalcoholic Fatty Liver: A Mini-ReviewDocument8 pagesReview Article: Signal Transductions and Nonalcoholic Fatty Liver: A Mini-ReviewFebyan AbotNo ratings yet
- Tumour Suppressor Genes: Jeffrey M Skolnik, David E FisherDocument6 pagesTumour Suppressor Genes: Jeffrey M Skolnik, David E FisherRicardo SilvaNo ratings yet
- Cancer-Related Inflammation, The Seventh Hallmark of Cancer: Links To Genetic InstabilityDocument9 pagesCancer-Related Inflammation, The Seventh Hallmark of Cancer: Links To Genetic Instabilitylucia coronaNo ratings yet
- NF-κB signalingDocument13 pagesNF-κB signalingAndri Praja SatriaNo ratings yet
- Cddis 2014201Document12 pagesCddis 2014201pedrolucs98No ratings yet
- Tnf-A Protein Synthesis Inhibitor Restores Neuronal Function and Reverses Cognitive Deficits Induced by Chronic NeuroinflammationDocument13 pagesTnf-A Protein Synthesis Inhibitor Restores Neuronal Function and Reverses Cognitive Deficits Induced by Chronic NeuroinflammationAlex AlexNo ratings yet
- Apoptosis and Therapy: Cold Spring Harbor Laboratory, Cold Spring Harbor, NY 11724, U.S.ADocument11 pagesApoptosis and Therapy: Cold Spring Harbor Laboratory, Cold Spring Harbor, NY 11724, U.S.AAntonio RolonNo ratings yet
- N Comms 7520Document12 pagesN Comms 7520kNo ratings yet
- Pi Is 1556086415330872Document2 pagesPi Is 1556086415330872Muthia FadhilaNo ratings yet
- Biology 12 00189 v3Document20 pagesBiology 12 00189 v3Albert FengNo ratings yet
- Impact of Genotype and Morphology On The Prognosis of GlioblastomaDocument8 pagesImpact of Genotype and Morphology On The Prognosis of GlioblastomaALINo ratings yet
- GB 16Document5 pagesGB 16ALINo ratings yet
- GB 2Document7 pagesGB 2ALINo ratings yet
- Prognostic Factors For Long-Term Survival After GlioblastomaDocument4 pagesPrognostic Factors For Long-Term Survival After GlioblastomaALINo ratings yet
- Upregulation of Mir-196B Confers A Poor Prognosis in Glioblastoma Patients Via Inducing A Proliferative PhenotypeDocument7 pagesUpregulation of Mir-196B Confers A Poor Prognosis in Glioblastoma Patients Via Inducing A Proliferative PhenotypeALINo ratings yet
- Original Article: High Expression of WDR1 in Primary Glioblastoma Is Associated With Poor PrognosisDocument12 pagesOriginal Article: High Expression of WDR1 in Primary Glioblastoma Is Associated With Poor PrognosisALINo ratings yet
- Predicting Glioblastoma Prognosis Networks Using Weighted Gene Co-Expression Network Analysis On TCGA DataDocument8 pagesPredicting Glioblastoma Prognosis Networks Using Weighted Gene Co-Expression Network Analysis On TCGA DataALINo ratings yet
- Surgical Neurology International: Socioeconomic Status Does Not Affect Prognosis in Patients With Glioblastoma MultiformeDocument9 pagesSurgical Neurology International: Socioeconomic Status Does Not Affect Prognosis in Patients With Glioblastoma MultiformeALINo ratings yet
- The Importance of Genomic Copy Number Changes in The Prognosis of Glioblastoma MultiformeDocument7 pagesThe Importance of Genomic Copy Number Changes in The Prognosis of Glioblastoma MultiformeALINo ratings yet
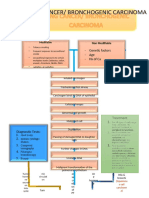
- Lung Cancer PathophysiologyDocument7 pagesLung Cancer PathophysiologyKristian Karl Bautista Kiw-isNo ratings yet
- Does Speculum Lubricant Affect Liquid Based Papanicolaou Test Adequacy?Document6 pagesDoes Speculum Lubricant Affect Liquid Based Papanicolaou Test Adequacy?Cristina CostaNo ratings yet
- MEDSURG - Cellular AberrationDocument10 pagesMEDSURG - Cellular AberrationLeslie CruzNo ratings yet
- 3 - NCM112Document6 pages3 - NCM112Edthel IniegoNo ratings yet
- Case Study Chapter 11Document2 pagesCase Study Chapter 11HugsNo ratings yet
- Cancer Incorporated Web Version PDFDocument247 pagesCancer Incorporated Web Version PDFDuoAmazonasNo ratings yet
- Assessing The Breasts and Regional LymphaticsDocument23 pagesAssessing The Breasts and Regional LymphaticsHannaNo ratings yet
- Steliarova Foucher - Et - Al 2005 CancerDocument11 pagesSteliarova Foucher - Et - Al 2005 CancerNur Melani Sari WardaniNo ratings yet
- Diagnostic Accuracy in The Detection of Depth of Myometrial Invasion With MRI in Early-Stage Endometrial CancerDocument5 pagesDiagnostic Accuracy in The Detection of Depth of Myometrial Invasion With MRI in Early-Stage Endometrial CancerBOHR International Journal on GynaecologyNo ratings yet
- BoneDocument98 pagesBoneElias Emmanuel JaimeNo ratings yet
- Colorectal Cancer: By:-Vaibhav SwarupDocument18 pagesColorectal Cancer: By:-Vaibhav SwarupOlga GoryachevaNo ratings yet
- Giant Cell Lesions of JawsDocument4 pagesGiant Cell Lesions of JawsRavish MalhotraNo ratings yet
- Clinico-Histopathological Analysis of Orbito-Ocular Lesions: A Hospital-Based StudyDocument6 pagesClinico-Histopathological Analysis of Orbito-Ocular Lesions: A Hospital-Based StudyFKUMP17 Part 2No ratings yet
- Ca72 4Document2 pagesCa72 4wongso100% (1)
- Soal Bhs Inggris Kelas 11 Wajib Ganjil 2022 - 2023Document12 pagesSoal Bhs Inggris Kelas 11 Wajib Ganjil 2022 - 2023SYUKRI, S.Pd.INo ratings yet
- RTOG 0529 IMRT Anal CancerDocument57 pagesRTOG 0529 IMRT Anal CancergammasharkNo ratings yet
- Dr. Rajasekhar A. National Pathology Laboratory HyderabadDocument26 pagesDr. Rajasekhar A. National Pathology Laboratory HyderabadAmit KumarNo ratings yet
- Breast CarcinomasDocument19 pagesBreast CarcinomasGirkati ShivaniNo ratings yet
- Guidelines For Primary HPV Testing For Cervical Cancer Screening in MalaysiaDocument38 pagesGuidelines For Primary HPV Testing For Cervical Cancer Screening in MalaysiadrnahafizahNo ratings yet
- Breast Cancer Review ArticleDocument1 pageBreast Cancer Review ArticleNazim ThakranNo ratings yet
- Community Health Survey: Assessment of Resident Health Needs and ConcernsDocument40 pagesCommunity Health Survey: Assessment of Resident Health Needs and ConcernsPreeti SachanNo ratings yet
- The Cure of CancerDocument2 pagesThe Cure of CancerLinh TrầnNo ratings yet
- American J Hematol - 2013 - Andersen - Eosinophilia in Routine Blood Samples and The Subsequent Risk of HematologicalDocument5 pagesAmerican J Hematol - 2013 - Andersen - Eosinophilia in Routine Blood Samples and The Subsequent Risk of HematologicalBelajarNo ratings yet
- Thom Lodovik Steviano Da Lopez - 19710004Document22 pagesThom Lodovik Steviano Da Lopez - 19710004Antonius BudionoNo ratings yet
- Block 5 (40) Female Reproductive System - Breast QsDocument42 pagesBlock 5 (40) Female Reproductive System - Breast QsJÓNATAN OSORIO PALACIONo ratings yet
- Mammograms: How Is A Mammogram Done?Document2 pagesMammograms: How Is A Mammogram Done?Mustafa FahamNo ratings yet
- Demystifying The Emf MythDocument56 pagesDemystifying The Emf Mythfrances leana capellan100% (1)
- Lateral Oncoplastic Breast Surgery (LOBS)Document5 pagesLateral Oncoplastic Breast Surgery (LOBS)Christian Daniel GarcíaNo ratings yet
- Update On Dermoscopy of SpitzReed Naevi and Management Guidelines by The International Dermoscopy SocietyDocument11 pagesUpdate On Dermoscopy of SpitzReed Naevi and Management Guidelines by The International Dermoscopy SocietyEduarda QuartinNo ratings yet