Download as pdf or txt
You might also like
- AACR 2022 Proceedings: Part A Online-Only and April 10From EverandAACR 2022 Proceedings: Part A Online-Only and April 10No ratings yet
- Cao 2011Document6 pagesCao 2011taoufik akabliNo ratings yet
- Methoxydalbergione Inhibits Esophageal Carcinoma Cell Proliferation and Migration by Inactivating NF‑ΚBDocument9 pagesMethoxydalbergione Inhibits Esophageal Carcinoma Cell Proliferation and Migration by Inactivating NF‑ΚBRepositorioNo ratings yet
- Biochemical and Biophysical Research CommunicationsDocument8 pagesBiochemical and Biophysical Research CommunicationsDelicz TanNo ratings yet
- Cannabidiol Inhibits Angiogenesis by Multiple Mechanisms PDFDocument14 pagesCannabidiol Inhibits Angiogenesis by Multiple Mechanisms PDFMartin SilvasNo ratings yet
- AZITROMICINADocument8 pagesAZITROMICINAEnrique MirandaNo ratings yet
- Fisetin Prevents Angiogenesis in Diabetic Retinopathy by Downregulating VEGFDocument7 pagesFisetin Prevents Angiogenesis in Diabetic Retinopathy by Downregulating VEGFvirginiaNo ratings yet
- Metabolic Reprogramming Due To Hypoxia in Pancreatic Can - 2021 - BiomedicineDocument14 pagesMetabolic Reprogramming Due To Hypoxia in Pancreatic Can - 2021 - BiomedicineAkhmad BajuriNo ratings yet
- Turmeric Is Therapeutic in Vivo On Patient-Derived Colorectal Cancer Xenografts: Inhibition of Growth, Metastasis, and Tumor RecurrenceDocument16 pagesTurmeric Is Therapeutic in Vivo On Patient-Derived Colorectal Cancer Xenografts: Inhibition of Growth, Metastasis, and Tumor RecurrenceFilipe GonçalvesNo ratings yet
- PEG-conjugated Hemoglobin Combination With Cisplatin Enforced The Antiangiogeic Effect in A Cervical Tumor Xenograft ModelDocument12 pagesPEG-conjugated Hemoglobin Combination With Cisplatin Enforced The Antiangiogeic Effect in A Cervical Tumor Xenograft ModelIstván PortörőNo ratings yet
- Gut Microbe Emediated Suppression of in Ammation-Associated Colon Carcinogenesis by Luminal Histamine ProductionDocument14 pagesGut Microbe Emediated Suppression of in Ammation-Associated Colon Carcinogenesis by Luminal Histamine ProductionNejc KovačNo ratings yet
- Effect of Artificial Oxygen Carrier With Chemotherapy On Tumor Hypoxia and NeovascularizationDocument9 pagesEffect of Artificial Oxygen Carrier With Chemotherapy On Tumor Hypoxia and NeovascularizationIstván PortörőNo ratings yet
- Redox BiologyDocument12 pagesRedox BiologyLin LINo ratings yet
- Li 2009Document10 pagesLi 2009bixagif369No ratings yet
- Inhibición de La Angiogénesis Tumoral Por CannabinoidesDocument16 pagesInhibición de La Angiogénesis Tumoral Por CannabinoidesjessicaNo ratings yet
- Abstract. Vascular Endothelial Growth Factor (VEGF) Report-: ONCOLOGY LETTERS 5: 1682-1686, 2013Document5 pagesAbstract. Vascular Endothelial Growth Factor (VEGF) Report-: ONCOLOGY LETTERS 5: 1682-1686, 2013yuliaNo ratings yet
- CXCR2 InhibitorDocument11 pagesCXCR2 InhibitorSaheed AbdulkarimNo ratings yet
- Thno 54961r2 3Document45 pagesThno 54961r2 3Eduardo JuarezNo ratings yet
- 0300060520925693Document11 pages0300060520925693Narjes Sharif AskariNo ratings yet
- UBR7 Inhibits HCC Tumorigenesis by Targeting Keap1/Nrf2/Bach1/HK2 and GlycolysisDocument18 pagesUBR7 Inhibits HCC Tumorigenesis by Targeting Keap1/Nrf2/Bach1/HK2 and GlycolysiskkurrriNo ratings yet
- 02 - VEGFR1-3 Axitinib Xenograft IJC2010Document11 pages02 - VEGFR1-3 Axitinib Xenograft IJC2010Omar SettiNo ratings yet
- Oncoimmune22v11p2054305 Cancer ICD HMGB1 Dendritic Very ImportDocument16 pagesOncoimmune22v11p2054305 Cancer ICD HMGB1 Dendritic Very Importichengmelody0108No ratings yet
- Accepted Manuscript: The International Journal of Biochemistry & Cell BiologyDocument37 pagesAccepted Manuscript: The International Journal of Biochemistry & Cell BiologyJuanA.UlloaNo ratings yet
- Ceju 69 00726Document9 pagesCeju 69 00726IkaNo ratings yet
- Metformin Inhibits Metastatic Breast Cancer Progression and Improves Chemosensitivity by Inducing Vessel Normalization Via PDGF-B DownregulationDocument17 pagesMetformin Inhibits Metastatic Breast Cancer Progression and Improves Chemosensitivity by Inducing Vessel Normalization Via PDGF-B DownregulationJackson HakimNo ratings yet
- Molecules 28 01352 With CoverDocument21 pagesMolecules 28 01352 With CoveracheikhrouhouNo ratings yet
- DNA Repair:EpigeneticsDocument20 pagesDNA Repair:EpigeneticssharonNo ratings yet
- Omcl2021 9068850Document31 pagesOmcl2021 9068850Heloisa PessoaNo ratings yet
- Oncolytic PresentationDocument10 pagesOncolytic PresentationIceyfloesNo ratings yet
- Fimmu 11 613114Document16 pagesFimmu 11 613114AquarelinhasNo ratings yet
- 05 ShivpriyaManchali PG 158 165ISpgrDocument18 pages05 ShivpriyaManchali PG 158 165ISpgrKarim HosniNo ratings yet
- Palizgir 2018Document7 pagesPalizgir 2018bharathbuddyNo ratings yet
- Biological Activity of Bevacizumab, A Humanized anti-VEGF Antibody in VitroDocument11 pagesBiological Activity of Bevacizumab, A Humanized anti-VEGF Antibody in VitroNguyễn Thị Yến NhiNo ratings yet
- Anticancer 7Document12 pagesAnticancer 7miral abibsiNo ratings yet
- Cell Cycle PaperDocument7 pagesCell Cycle Paperapi-136834987No ratings yet
- AtomicBomb Scid MiceDocument7 pagesAtomicBomb Scid MiceDavid MartínezNo ratings yet
- Transplant International Special Issue: Abstracts of The 14th Congress of The European Society For Organ Transplantation Volume 22, Issue Supplement s2, Pages 95-222, August 2009Document128 pagesTransplant International Special Issue: Abstracts of The 14th Congress of The European Society For Organ Transplantation Volume 22, Issue Supplement s2, Pages 95-222, August 2009Enrique Moreno GonzálezNo ratings yet
- ReishiMax Clinical StudyDocument7 pagesReishiMax Clinical StudynsagelocspaNo ratings yet
- Clostridium Novyi-Nt in Cancer Therapy: SciencedirectDocument9 pagesClostridium Novyi-Nt in Cancer Therapy: SciencedirectAde Putra RizkyNo ratings yet
- 1 s2.0 S0753332222011520 MainDocument20 pages1 s2.0 S0753332222011520 MainTejas krishnakanthNo ratings yet
- Mdpi2 AntioxidantsDocument14 pagesMdpi2 AntioxidantsKatarinaNo ratings yet
- s12885 015 1137 9Document13 pagess12885 015 1137 9ahmad saadNo ratings yet
- Cholecystokinin Receptor Antagonist Improves Efficacy of Chemotherapy in Murine Models of Pancreatic Cancer by Altering The Tumor MicroenvironmentDocument21 pagesCholecystokinin Receptor Antagonist Improves Efficacy of Chemotherapy in Murine Models of Pancreatic Cancer by Altering The Tumor MicroenvironmentJaya Kunal DubeyNo ratings yet
- 1247 FTPDocument10 pages1247 FTPMeow WeddingNo ratings yet
- Intra-Lesional Injection of The Novel PKC Activator EBC-46 Rapidly Ablates Tumors in Mouse ModelsDocument12 pagesIntra-Lesional Injection of The Novel PKC Activator EBC-46 Rapidly Ablates Tumors in Mouse ModelscarlammartinsNo ratings yet
- Ultrasound-Targeted Microbubble Destruction Mediated Herpes Simplex Virus-Thymidine Kinase Gene Treats Hepatoma in MiceDocument6 pagesUltrasound-Targeted Microbubble Destruction Mediated Herpes Simplex Virus-Thymidine Kinase Gene Treats Hepatoma in MiceIndra17No ratings yet
- 10 1016@j Biopha 2019 109142Document13 pages10 1016@j Biopha 2019 109142CoNo ratings yet
- 2023 Int J Mol Sci 10581 (Nagy)Document17 pages2023 Int J Mol Sci 10581 (Nagy)Lázár LászlóNo ratings yet
- (In Vivo) 6823520Document11 pages(In Vivo) 6823520ingelitaNo ratings yet
- Shengbai Decoction Enhances The Anti Tumor Efficacy of - 2021 - Biomedicine - P PDFDocument10 pagesShengbai Decoction Enhances The Anti Tumor Efficacy of - 2021 - Biomedicine - P PDFFernando MaffeisNo ratings yet
- INCLUIDOS-clusianone and Cancer - Search Results - PubMedDocument6 pagesINCLUIDOS-clusianone and Cancer - Search Results - PubMedLidiaAmorimNo ratings yet
- Effect of Platelet-Derived Growth Factor-B On Renal Cell Carcinoma Growth and ProgressionDocument11 pagesEffect of Platelet-Derived Growth Factor-B On Renal Cell Carcinoma Growth and ProgressionOZGE DENIZ YESILNo ratings yet
- Kjuo009 02 03Document7 pagesKjuo009 02 03Gambit Marvel FanNo ratings yet
- Anticancer Activity of Phyllanthus Niruri Linn Extract in Colorectal Cancer Patients A Phase II Clinical TrialDocument5 pagesAnticancer Activity of Phyllanthus Niruri Linn Extract in Colorectal Cancer Patients A Phase II Clinical TrialAndi SaputraNo ratings yet
- Lister Strain Vaccinia Virus, A Potential Therapeutic Vector Targeting Hypoxic TumoursDocument15 pagesLister Strain Vaccinia Virus, A Potential Therapeutic Vector Targeting Hypoxic TumourssusasureshNo ratings yet
- 2022 01 07 475451v1 FullDocument35 pages2022 01 07 475451v1 Fulljongsu kimNo ratings yet
- Sulforaphane Regulates Self-Renewal of Pancreatic Cancer Stem Cells Through The Modulation of Sonic hedgehog-GLI PathwayDocument11 pagesSulforaphane Regulates Self-Renewal of Pancreatic Cancer Stem Cells Through The Modulation of Sonic hedgehog-GLI PathwayMikhail PisarevNo ratings yet
- Circulating Endothelial Cells and Other Angiogenesis Factors in Pancreatic Carcinoma Patients Receiving Gemcitabine ChemotherapyDocument10 pagesCirculating Endothelial Cells and Other Angiogenesis Factors in Pancreatic Carcinoma Patients Receiving Gemcitabine ChemotherapyftwNo ratings yet
- AACR 2017 Proceedings: Abstracts 1-3062From EverandAACR 2017 Proceedings: Abstracts 1-3062No ratings yet
- AACR 2017 Proceedings: Abstracts 3063-5947From EverandAACR 2017 Proceedings: Abstracts 3063-5947No ratings yet
- Association Between Vegf 936cT Gene Polymorphism With Gastric Premalignant Lesions and Vegf Levels in Helicobacter Pylori PatientsDocument3 pagesAssociation Between Vegf 936cT Gene Polymorphism With Gastric Premalignant Lesions and Vegf Levels in Helicobacter Pylori PatientsDavid NicholasNo ratings yet
- Rev of Pat and Gen 7 TH EdiDocument707 pagesRev of Pat and Gen 7 TH EdihatsuneNo ratings yet
- POTENZA - Radiofrequency Irradiation Attenuates Angiogenesis and Infl... - I...Document20 pagesPOTENZA - Radiofrequency Irradiation Attenuates Angiogenesis and Infl... - I...TimNo ratings yet
- VEGFDocument20 pagesVEGFLivia Rhea AlvitaNo ratings yet
- Cryopreservation of Skin TissuesDocument21 pagesCryopreservation of Skin TissuesArchanaShenoyNo ratings yet
- Public Health Notice - Long COVIDDocument2 pagesPublic Health Notice - Long COVIDAnthony DocKek PenaNo ratings yet
- Wounds Gen PathDocument26 pagesWounds Gen PathCynthia CNo ratings yet
- Anti-VEGF Agents and Their Clinical Applications: Dr. Sriniwas Atal MD. Resident OphthalmologyDocument67 pagesAnti-VEGF Agents and Their Clinical Applications: Dr. Sriniwas Atal MD. Resident OphthalmologySriniwasNo ratings yet
- Chronic Obstructive Pulmonary Disease and The Cardiovascular System Vascular Repair and Regeneration As A Therapeutic TargetDocument26 pagesChronic Obstructive Pulmonary Disease and The Cardiovascular System Vascular Repair and Regeneration As A Therapeutic TargetAndreea MoalesNo ratings yet
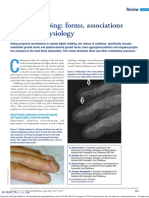
- Digital ClubbingDocument6 pagesDigital ClubbingPedro AzevedoNo ratings yet
- A Comparitive and Prognostic Study of Circulating Tumor Cells and Vegfr-2 in Different Histological Grades of Oral Squamous Cell CarcinomaDocument10 pagesA Comparitive and Prognostic Study of Circulating Tumor Cells and Vegfr-2 in Different Histological Grades of Oral Squamous Cell CarcinomaIJAR JOURNALNo ratings yet
- 1 s2.0 S2096691122000723 MainDocument7 pages1 s2.0 S2096691122000723 MainMaya IdayahNo ratings yet
- Estudio CarmenaDocument11 pagesEstudio CarmenaAlfredo BalcázarNo ratings yet
- Cancer Prevention - From Mechanisms To Translational BenefitsDocument488 pagesCancer Prevention - From Mechanisms To Translational BenefitsJosé RamírezNo ratings yet
- Rs3pe SyndromeDocument6 pagesRs3pe SyndromejhonnyNo ratings yet
- Lipedema An Overview of Its Clinical ManifestationsDocument10 pagesLipedema An Overview of Its Clinical ManifestationsCarla a s MoreiraNo ratings yet
- 21 Natural Scientific Pathways Shown To Stimulate Hair Growth and Prevent Hair Loss 21 Natural ... (PDFDrive)Document36 pages21 Natural Scientific Pathways Shown To Stimulate Hair Growth and Prevent Hair Loss 21 Natural ... (PDFDrive)Mohamed KhaterNo ratings yet
- Artigo 8Document10 pagesArtigo 8daisyscholliersNo ratings yet
- Tumor Angiogenesis: Causes, Consequences, Challenges and OpportunitiesDocument26 pagesTumor Angiogenesis: Causes, Consequences, Challenges and OpportunitiesStella NoviaNo ratings yet
- NatureCeuticals 2020 ProspectusDocument10 pagesNatureCeuticals 2020 ProspectusJamall EllisNo ratings yet
- Tinjauan Manfaat Bunga Telang (Clitoria Ternatea L.) Bagi Kesehatan ManusiaDocument23 pagesTinjauan Manfaat Bunga Telang (Clitoria Ternatea L.) Bagi Kesehatan Manusiakurnia saptaNo ratings yet
- Hallmarks of Cancer: The Next Generation: ReviewDocument29 pagesHallmarks of Cancer: The Next Generation: Review122307403No ratings yet
- Low-Level Laser Therapy (780 NM) On VEGF Modulation at Partially Injured Achilles TendonDocument5 pagesLow-Level Laser Therapy (780 NM) On VEGF Modulation at Partially Injured Achilles TendonMichele GonçalvesNo ratings yet
- For Peer Review OnlyDocument45 pagesFor Peer Review Onlyros_galindoNo ratings yet
- Pterigium PDFDocument152 pagesPterigium PDFArizkamhNo ratings yet
- Benzimidazole - A Medicinally Important Heterocyclic MoietyDocument15 pagesBenzimidazole - A Medicinally Important Heterocyclic MoietyKeyvanNo ratings yet
- Ultrasound in The Prevention, Diagnosis and Management of Ovarian Hyperstimulation Syndrome in BamakoDocument9 pagesUltrasound in The Prevention, Diagnosis and Management of Ovarian Hyperstimulation Syndrome in BamakoIJAR JOURNALNo ratings yet
- Polymeric Nanoparticles For Cancer Therapy: Suphiya Parveen & Sanjeeb K. SahooDocument16 pagesPolymeric Nanoparticles For Cancer Therapy: Suphiya Parveen & Sanjeeb K. SahooFaizhal Dimas LeksonoNo ratings yet
- Radbio 2010 Study GuideDocument194 pagesRadbio 2010 Study GuideDade212No ratings yet
- J Hepato Biliary Pancreat - 2014 - Aishima - Pathogenesis and Classification of Intrahepatic Cholangiocarcinoma DifferentDocument7 pagesJ Hepato Biliary Pancreat - 2014 - Aishima - Pathogenesis and Classification of Intrahepatic Cholangiocarcinoma DifferentleartaNo ratings yet