
The Epididymis Re-Visited: A Personal View: Experience and History
The Epididymis Re-Visited: A Personal View: Experience and History
You might also like
- Anatomy of The Reproductive System EDocument4 pagesAnatomy of The Reproductive System EshreeNo ratings yet
- 2002 - The Role of Colesterol Efflux in Regulating The Fertilization Potential of Mammalian SpermatozoaDocument7 pages2002 - The Role of Colesterol Efflux in Regulating The Fertilization Potential of Mammalian Spermatozoadavid torresNo ratings yet
- (17417899 - Reproduction) Epididymosomes, Prostasomes, and Liposomes - Their Roles in Mammalian Male Reproductive PhysiologyDocument15 pages(17417899 - Reproduction) Epididymosomes, Prostasomes, and Liposomes - Their Roles in Mammalian Male Reproductive PhysiologyAhmad SolihinNo ratings yet
- Sperm Proteome Maturation in The Mouse EpididymisDocument23 pagesSperm Proteome Maturation in The Mouse EpididymisAhmad SolihinNo ratings yet
- Fertilization: Recommended Reading: SummaryDocument10 pagesFertilization: Recommended Reading: SummaryKnjigeNo ratings yet
- 3.3 Fertilization in Plants and AnimalsDocument56 pages3.3 Fertilization in Plants and AnimalsAhmed AimanNo ratings yet
- Fertilizacion RevisionDocument8 pagesFertilizacion RevisionAsistente IRP RESTREPONo ratings yet
- Biolreprod 0247Document9 pagesBiolreprod 0247lucascotaNo ratings yet
- Mechanisms of Mammalian Sperm Egg Interaction Leading To Fertilization 2161 0932.1000e107Document4 pagesMechanisms of Mammalian Sperm Egg Interaction Leading To Fertilization 2161 0932.1000e1071005 Siti Ainunnisa Lingga WinariNo ratings yet
- Cib 6 E26729Document3 pagesCib 6 E26729Kaio FernandesNo ratings yet
- Legends of The Sperm and EggDocument8 pagesLegends of The Sperm and EggALNo ratings yet
- Pudney 1995 PDFDocument39 pagesPudney 1995 PDFKei MlNo ratings yet
- @artigo Novo 2Document6 pages@artigo Novo 2Beatrice MacenteNo ratings yet
- Great Sperm RaceDocument19 pagesGreat Sperm RacefrjesNo ratings yet
- Human FertilizationDocument3 pagesHuman Fertilizationtamalbio7No ratings yet
- Biological Basis For Human Capacitation: Christopher de JongeDocument10 pagesBiological Basis For Human Capacitation: Christopher de JongeLara RamundoNo ratings yet
- Biological Basis For Human Capacitation: Christopher de JongeDocument10 pagesBiological Basis For Human Capacitation: Christopher de JongeVicNo ratings yet
- ArticuloDocument16 pagesArticuloMiguel Ángel Carmona ZamudioNo ratings yet
- Human Sperm Capacitation and The Acrosome Reaction : L.J.D.Zaneveld, C.J.De Jonge, R.A.Anderson and S.R.MackDocument10 pagesHuman Sperm Capacitation and The Acrosome Reaction : L.J.D.Zaneveld, C.J.De Jonge, R.A.Anderson and S.R.MackDaniel LunaNo ratings yet
- Sperm PreparationDocument10 pagesSperm Preparationcarlosdanielrc1983No ratings yet
- Fertilization: Beginning A New Organism: Lee Kui SoonDocument31 pagesFertilization: Beginning A New Organism: Lee Kui SoonSayapPutihNo ratings yet
- Biology: Puzzles of Mammalian Fertilization - and BeyondDocument13 pagesBiology: Puzzles of Mammalian Fertilization - and BeyondKaren Petite Rubio MoralesNo ratings yet
- Review of Prostate Anatomy and Embryology and The Etiology of Benign Prostatic Hyperplasia PDFDocument10 pagesReview of Prostate Anatomy and Embryology and The Etiology of Benign Prostatic Hyperplasia PDFAlejandro GuerreroNo ratings yet
- FertilizationDocument11 pagesFertilization137 - ShubhamNo ratings yet
- Biology: Find and Fuse: Unsolved Mysteries in Sperm - Egg RecognitionDocument11 pagesBiology: Find and Fuse: Unsolved Mysteries in Sperm - Egg RecognitionFlavio NavarreteNo ratings yet
- 1 s2.0 S0093691X13003282 MainDocument7 pages1 s2.0 S0093691X13003282 Maincarrerajm22No ratings yet
- Vmi2011 757194Document13 pagesVmi2011 757194Ahmad SolihinNo ratings yet
- Protein Signatures of Seminal Plasma From Bulls With Contrasting Frozen-Thawed Sperm ViabilityDocument14 pagesProtein Signatures of Seminal Plasma From Bulls With Contrasting Frozen-Thawed Sperm Viabilitysaifulmangopo123No ratings yet
- Paternal Investment and Lntracellular Sperm-Egg Interactions During and Following Fer T Zat On inDocument27 pagesPaternal Investment and Lntracellular Sperm-Egg Interactions During and Following Fer T Zat On inじょしら フィアンナNo ratings yet
- Alakeshbarman Zoology PG 2nd FertilizationDocument28 pagesAlakeshbarman Zoology PG 2nd Fertilizationouafia liiamaniNo ratings yet
- Topping Off A Mechanism of First - Males PrecedenceDocument4 pagesTopping Off A Mechanism of First - Males PrecedenceJavier Aldo BianchiNo ratings yet
- Moriyama 2018Document8 pagesMoriyama 2018Gabriel BuonoNo ratings yet
- Quality of Canine Spermatozoa RetrievedDocument5 pagesQuality of Canine Spermatozoa Retrievedİhsan DoğuNo ratings yet
- Description of Reproductive System of Indian Water Scorpion, Laccotrephes Maculatus Fabr. (Hemiptera, Heteroptera: Nepidae)Document13 pagesDescription of Reproductive System of Indian Water Scorpion, Laccotrephes Maculatus Fabr. (Hemiptera, Heteroptera: Nepidae)Kanhiya MahourNo ratings yet
- AlakeshBarman Zoology PG 2ND FERTILIZATIONDocument28 pagesAlakeshBarman Zoology PG 2ND FERTILIZATIONTrueAlpha 16No ratings yet
- FertilizationDocument10 pagesFertilizationdeepakshi rajputNo ratings yet
- The Mouse Epididymal Transcriptome: Transcriptional Profiling of Segmental Gene Expression in The EpididymisDocument10 pagesThe Mouse Epididymal Transcriptome: Transcriptional Profiling of Segmental Gene Expression in The EpididymisAhmad SolihinNo ratings yet
- Reproduction PPT 6 PHYSIOLOGY OF PREGNANCYDocument98 pagesReproduction PPT 6 PHYSIOLOGY OF PREGNANCYlisanames.23No ratings yet
- Human Development Days 1-4Document43 pagesHuman Development Days 1-4Kriztine Mae GonzalesNo ratings yet
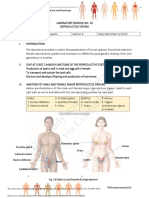
- Lab Exercise19 Reproductive System PDFDocument3 pagesLab Exercise19 Reproductive System PDFcsolutionNo ratings yet
- Asc101 Lab Exercise No. 3Document10 pagesAsc101 Lab Exercise No. 3MELITO JR. CATAYLONo ratings yet
- Developmental Biology: GametogenesisDocument21 pagesDevelopmental Biology: GametogenesisTapan Kumar PalNo ratings yet
- Marine Biological Laboratory Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Biological BulletinDocument21 pagesMarine Biological Laboratory Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Biological BulletinRayara Matos Fontineles GuedesNo ratings yet
- 6i. Male Reproduction - OnPRC Module 2-Student HandoutDocument12 pages6i. Male Reproduction - OnPRC Module 2-Student HandoutAkash ShawNo ratings yet
- Journal of Morphology - 2020 - Starck - Morphology of The Avian Yolk SacDocument14 pagesJournal of Morphology - 2020 - Starck - Morphology of The Avian Yolk SacMashai LesenyehoNo ratings yet
- Cortical Granule FunctionDocument17 pagesCortical Granule FunctionEric PhillipsNo ratings yet
- Science Aax7890 PDFDocument9 pagesScience Aax7890 PDFAndoingNo ratings yet
- Acrosome ReactionDocument4 pagesAcrosome ReactionKuldip DwivediNo ratings yet
- 1999 Ency HessDocument8 pages1999 Ency HessKishan GaurNo ratings yet
- Role of Glycolysis and Respiration in Sperm Metabolism and MotilityDocument60 pagesRole of Glycolysis and Respiration in Sperm Metabolism and MotilitywalczakcNo ratings yet
- Fertilization Overview: Sperm PenetrationDocument13 pagesFertilization Overview: Sperm PenetrationAmira FahruddinNo ratings yet
- Biology - Human ReproductionDocument21 pagesBiology - Human Reproductionsgw67No ratings yet
- LAB EXERCISE 2 - OogenesisDocument4 pagesLAB EXERCISE 2 - OogenesisGerald Angelo DeguinioNo ratings yet
- Biology of Polyspermy in IVF and Its Clinical IndicationDocument6 pagesBiology of Polyspermy in IVF and Its Clinical IndicationMD LarasatiNo ratings yet
- Transcriptional Profiling and Physiological Roles of Aedes Aegypti Spermathecal-Related GenesDocument18 pagesTranscriptional Profiling and Physiological Roles of Aedes Aegypti Spermathecal-Related GenesSara VillaNo ratings yet
- tmpD7B9 TMPDocument12 pagestmpD7B9 TMPFrontiersNo ratings yet
- (Unit 4) Human GametesDocument8 pages(Unit 4) Human GametesHanif AlfarizkiNo ratings yet
- Bao Quan Mo Tinh HoanDocument21 pagesBao Quan Mo Tinh Hoanhhhhhhhhhh0000No ratings yet
- Lab Report 4 GametogenesisDocument13 pagesLab Report 4 GametogenesisWnz NaiveNo ratings yet
- Investigating Reproducibility and Tracking ProvenaDocument15 pagesInvestigating Reproducibility and Tracking ProvenaAhmad SolihinNo ratings yet
- Best Practices For Variant Calling in Clinical Sequencing: Review Open AccessDocument13 pagesBest Practices For Variant Calling in Clinical Sequencing: Review Open AccessAhmad SolihinNo ratings yet
- Assessment of The Genomic Variation in A Cattle Population by Re-Sequencing of Key Animals at Low To Medium CoverageDocument9 pagesAssessment of The Genomic Variation in A Cattle Population by Re-Sequencing of Key Animals at Low To Medium CoverageAhmad SolihinNo ratings yet
- ID Pengaruh Propylthiuouracil Terhadap KadaDocument8 pagesID Pengaruh Propylthiuouracil Terhadap KadaAhmad SolihinNo ratings yet
- 1 s2.0 S0300483X20301670 MainDocument12 pages1 s2.0 S0300483X20301670 MainAhmad SolihinNo ratings yet
- Steroids: SciencedirectDocument11 pagesSteroids: SciencedirectAhmad SolihinNo ratings yet
- T-ARMS PCR Genotyping of SNP rs445709131 Using Thermostable Strand Displacement PolymeraseDocument5 pagesT-ARMS PCR Genotyping of SNP rs445709131 Using Thermostable Strand Displacement PolymeraseAhmad SolihinNo ratings yet
- Environmental Research: Yingying Yang, Luqing Pan, Yueyao Zhou, Ruiyi Xu, Dongyu LiDocument12 pagesEnvironmental Research: Yingying Yang, Luqing Pan, Yueyao Zhou, Ruiyi Xu, Dongyu LiAhmad SolihinNo ratings yet
- Guide To The Care and Use of Experimental Animals, Volume 2 (1984) Ix. DogsDocument14 pagesGuide To The Care and Use of Experimental Animals, Volume 2 (1984) Ix. DogsAhmad SolihinNo ratings yet
- Food and Chemical ToxicologyDocument10 pagesFood and Chemical ToxicologyAhmad SolihinNo ratings yet
- Testicular Biopsy: Clinical Practice and Interpretation: ReviewDocument6 pagesTesticular Biopsy: Clinical Practice and Interpretation: ReviewAhmad SolihinNo ratings yet
- The Maturation of Murine Spermatozoa Membranes Within The Epididymis A Computational Biology PerspectiveDocument11 pagesThe Maturation of Murine Spermatozoa Membranes Within The Epididymis A Computational Biology PerspectiveAhmad SolihinNo ratings yet
- Sperm Proteome Maturation in The Mouse EpididymisDocument23 pagesSperm Proteome Maturation in The Mouse EpididymisAhmad SolihinNo ratings yet
- Improving Bioscience Research Reporting: The ARRIVE Guidelines For Reporting Animal ResearchDocument5 pagesImproving Bioscience Research Reporting: The ARRIVE Guidelines For Reporting Animal ResearchAhmad SolihinNo ratings yet
- Molecular Biology of Spermatogenesis: Novel Targets of Apparently Idiopathic Male InfertilityDocument13 pagesMolecular Biology of Spermatogenesis: Novel Targets of Apparently Idiopathic Male InfertilityAhmad SolihinNo ratings yet
- Atlas Anatomi Dan Histologi Binatang Part 1Document15 pagesAtlas Anatomi Dan Histologi Binatang Part 1Ahmad SolihinNo ratings yet
- Annals Oi Anatomy - : Apocrine Secretion - Fact or Artifact?Document12 pagesAnnals Oi Anatomy - : Apocrine Secretion - Fact or Artifact?Ahmad SolihinNo ratings yet
- Pi Is 0021925820827409Document8 pagesPi Is 0021925820827409Ahmad SolihinNo ratings yet
- Vmi2011 757194Document13 pagesVmi2011 757194Ahmad SolihinNo ratings yet
- Practica Reproducitve Systme and PresentDocument10 pagesPractica Reproducitve Systme and PresentLauris CassoNo ratings yet
- PennnyssDocument33 pagesPennnyssscribdmefaischierNo ratings yet
- Male Reproductive SystemDocument15 pagesMale Reproductive SystemCharlz ZipaganNo ratings yet
- Physiologi of Reproductive System With Voice - CompressedDocument16 pagesPhysiologi of Reproductive System With Voice - CompressedMsatriaNo ratings yet
- S10 Q3 WEEK1 Reproductive System LECTUREDocument35 pagesS10 Q3 WEEK1 Reproductive System LECTUREREGLOS, Marie Nhelle K.No ratings yet
- Phimosis Cure TestimonyDocument3 pagesPhimosis Cure TestimonyEnrique RojasNo ratings yet
- Seminal Fluid AnalysisDocument11 pagesSeminal Fluid AnalysisKenneth Jake BatiduanNo ratings yet
- ARTIKEL ILMIAH (061611535039 - Novi Fitria Hanifah)Document10 pagesARTIKEL ILMIAH (061611535039 - Novi Fitria Hanifah)novi fitriaNo ratings yet
- Third Assignment (Forensic)Document2 pagesThird Assignment (Forensic)Dean Mark AnacioNo ratings yet
- Anatomy of The Male Reproductive SystemDocument70 pagesAnatomy of The Male Reproductive SystemDaniel HikaNo ratings yet
- 02 Overview of The Male and Female Reproductive SystemsDocument30 pages02 Overview of The Male and Female Reproductive SystemsOsman Saidu SesayNo ratings yet
- Ovozoa Vol. 7, No. 2, Oktober 2018 ISSN: 2302-6464Document6 pagesOvozoa Vol. 7, No. 2, Oktober 2018 ISSN: 2302-6464Miftah LailiNo ratings yet
- O'Hara & O'Hara - The Effect of Male Circumcision On The Sexual Enjoyment of The Female Partner PDFDocument6 pagesO'Hara & O'Hara - The Effect of Male Circumcision On The Sexual Enjoyment of The Female Partner PDFOmar MohammedNo ratings yet
- Anatomy For Beginners - Reproduction WorksheetDocument4 pagesAnatomy For Beginners - Reproduction WorksheetJames Dauray100% (1)
- Pourusha VastiDocument54 pagesPourusha Vastiksr prasadNo ratings yet
- The PenisDocument6 pagesThe Penisntambik21No ratings yet
- Pengaruh Suhu Dan Lama Thawing Di Dataran Rendah Terhadap Kualitas Semen Beku Sapi BrahmanDocument7 pagesPengaruh Suhu Dan Lama Thawing Di Dataran Rendah Terhadap Kualitas Semen Beku Sapi BrahmanEnikaaNo ratings yet
- Kelly9e Ch03 Male Sexual-Anatomy and PhysiologyDocument19 pagesKelly9e Ch03 Male Sexual-Anatomy and Physiologysauloalberto6254No ratings yet
- Histology Male ReproductiveDocument31 pagesHistology Male Reproductivekhiks34No ratings yet
- Be My Temptation (COMPLETED) - Shaleh - Wattpad - WattpadDocument256 pagesBe My Temptation (COMPLETED) - Shaleh - Wattpad - WattpadWeef hoofNo ratings yet
- 2.1.1 Reproductive Systems Teacher Answer SheetDocument3 pages2.1.1 Reproductive Systems Teacher Answer SheetFerdi Hasan100% (1)
- SpermDocument1 pageSpermEve Karen PobleteNo ratings yet
- Abordul Laparoscopic Pentru Testicule Nepalpabile Aspecte Tehnice Si RezultateDocument6 pagesAbordul Laparoscopic Pentru Testicule Nepalpabile Aspecte Tehnice Si RezultateAlexandra NechiforNo ratings yet
- Sperm Maturation in The Domestic CatDocument11 pagesSperm Maturation in The Domestic CatGeovanni LunaNo ratings yet
- Histologi Reproduksi PriaDocument45 pagesHistologi Reproduksi Priablack junoNo ratings yet
- Maximum Sex! by Editors of MaximDocument200 pagesMaximum Sex! by Editors of MaximLoto760% (3)
- Anatomy of The Male Reproductive SystemDocument24 pagesAnatomy of The Male Reproductive SystemAbbasi KahumaNo ratings yet
- Male Reproductive SystemDocument10 pagesMale Reproductive SystemTracy zorcaNo ratings yet
- Paraphimosis (Ingles)Document1 pageParaphimosis (Ingles)Joaquín SosaNo ratings yet
- Developmental Process of SpermatogenesisDocument4 pagesDevelopmental Process of Spermatogenesispricilia ardianiNo ratings yet









































































































Download as pdf or txt
You might also like
- Anatomy of The Reproductive System EDocument4 pagesAnatomy of The Reproductive System EshreeNo ratings yet
- 2002 - The Role of Colesterol Efflux in Regulating The Fertilization Potential of Mammalian SpermatozoaDocument7 pages2002 - The Role of Colesterol Efflux in Regulating The Fertilization Potential of Mammalian Spermatozoadavid torresNo ratings yet
- (17417899 - Reproduction) Epididymosomes, Prostasomes, and Liposomes - Their Roles in Mammalian Male Reproductive PhysiologyDocument15 pages(17417899 - Reproduction) Epididymosomes, Prostasomes, and Liposomes - Their Roles in Mammalian Male Reproductive PhysiologyAhmad SolihinNo ratings yet
- Sperm Proteome Maturation in The Mouse EpididymisDocument23 pagesSperm Proteome Maturation in The Mouse EpididymisAhmad SolihinNo ratings yet
- Fertilization: Recommended Reading: SummaryDocument10 pagesFertilization: Recommended Reading: SummaryKnjigeNo ratings yet
- 3.3 Fertilization in Plants and AnimalsDocument56 pages3.3 Fertilization in Plants and AnimalsAhmed AimanNo ratings yet
- Fertilizacion RevisionDocument8 pagesFertilizacion RevisionAsistente IRP RESTREPONo ratings yet
- Biolreprod 0247Document9 pagesBiolreprod 0247lucascotaNo ratings yet
- Mechanisms of Mammalian Sperm Egg Interaction Leading To Fertilization 2161 0932.1000e107Document4 pagesMechanisms of Mammalian Sperm Egg Interaction Leading To Fertilization 2161 0932.1000e1071005 Siti Ainunnisa Lingga WinariNo ratings yet
- Cib 6 E26729Document3 pagesCib 6 E26729Kaio FernandesNo ratings yet
- Legends of The Sperm and EggDocument8 pagesLegends of The Sperm and EggALNo ratings yet
- Pudney 1995 PDFDocument39 pagesPudney 1995 PDFKei MlNo ratings yet
- @artigo Novo 2Document6 pages@artigo Novo 2Beatrice MacenteNo ratings yet
- Great Sperm RaceDocument19 pagesGreat Sperm RacefrjesNo ratings yet
- Human FertilizationDocument3 pagesHuman Fertilizationtamalbio7No ratings yet
- Biological Basis For Human Capacitation: Christopher de JongeDocument10 pagesBiological Basis For Human Capacitation: Christopher de JongeLara RamundoNo ratings yet
- Biological Basis For Human Capacitation: Christopher de JongeDocument10 pagesBiological Basis For Human Capacitation: Christopher de JongeVicNo ratings yet
- ArticuloDocument16 pagesArticuloMiguel Ángel Carmona ZamudioNo ratings yet
- Human Sperm Capacitation and The Acrosome Reaction : L.J.D.Zaneveld, C.J.De Jonge, R.A.Anderson and S.R.MackDocument10 pagesHuman Sperm Capacitation and The Acrosome Reaction : L.J.D.Zaneveld, C.J.De Jonge, R.A.Anderson and S.R.MackDaniel LunaNo ratings yet
- Sperm PreparationDocument10 pagesSperm Preparationcarlosdanielrc1983No ratings yet
- Fertilization: Beginning A New Organism: Lee Kui SoonDocument31 pagesFertilization: Beginning A New Organism: Lee Kui SoonSayapPutihNo ratings yet
- Biology: Puzzles of Mammalian Fertilization - and BeyondDocument13 pagesBiology: Puzzles of Mammalian Fertilization - and BeyondKaren Petite Rubio MoralesNo ratings yet
- Review of Prostate Anatomy and Embryology and The Etiology of Benign Prostatic Hyperplasia PDFDocument10 pagesReview of Prostate Anatomy and Embryology and The Etiology of Benign Prostatic Hyperplasia PDFAlejandro GuerreroNo ratings yet
- FertilizationDocument11 pagesFertilization137 - ShubhamNo ratings yet
- Biology: Find and Fuse: Unsolved Mysteries in Sperm - Egg RecognitionDocument11 pagesBiology: Find and Fuse: Unsolved Mysteries in Sperm - Egg RecognitionFlavio NavarreteNo ratings yet
- 1 s2.0 S0093691X13003282 MainDocument7 pages1 s2.0 S0093691X13003282 Maincarrerajm22No ratings yet
- Vmi2011 757194Document13 pagesVmi2011 757194Ahmad SolihinNo ratings yet
- Protein Signatures of Seminal Plasma From Bulls With Contrasting Frozen-Thawed Sperm ViabilityDocument14 pagesProtein Signatures of Seminal Plasma From Bulls With Contrasting Frozen-Thawed Sperm Viabilitysaifulmangopo123No ratings yet
- Paternal Investment and Lntracellular Sperm-Egg Interactions During and Following Fer T Zat On inDocument27 pagesPaternal Investment and Lntracellular Sperm-Egg Interactions During and Following Fer T Zat On inじょしら フィアンナNo ratings yet
- Alakeshbarman Zoology PG 2nd FertilizationDocument28 pagesAlakeshbarman Zoology PG 2nd Fertilizationouafia liiamaniNo ratings yet
- Topping Off A Mechanism of First - Males PrecedenceDocument4 pagesTopping Off A Mechanism of First - Males PrecedenceJavier Aldo BianchiNo ratings yet
- Moriyama 2018Document8 pagesMoriyama 2018Gabriel BuonoNo ratings yet
- Quality of Canine Spermatozoa RetrievedDocument5 pagesQuality of Canine Spermatozoa Retrievedİhsan DoğuNo ratings yet
- Description of Reproductive System of Indian Water Scorpion, Laccotrephes Maculatus Fabr. (Hemiptera, Heteroptera: Nepidae)Document13 pagesDescription of Reproductive System of Indian Water Scorpion, Laccotrephes Maculatus Fabr. (Hemiptera, Heteroptera: Nepidae)Kanhiya MahourNo ratings yet
- AlakeshBarman Zoology PG 2ND FERTILIZATIONDocument28 pagesAlakeshBarman Zoology PG 2ND FERTILIZATIONTrueAlpha 16No ratings yet
- FertilizationDocument10 pagesFertilizationdeepakshi rajputNo ratings yet
- The Mouse Epididymal Transcriptome: Transcriptional Profiling of Segmental Gene Expression in The EpididymisDocument10 pagesThe Mouse Epididymal Transcriptome: Transcriptional Profiling of Segmental Gene Expression in The EpididymisAhmad SolihinNo ratings yet
- Reproduction PPT 6 PHYSIOLOGY OF PREGNANCYDocument98 pagesReproduction PPT 6 PHYSIOLOGY OF PREGNANCYlisanames.23No ratings yet
- Human Development Days 1-4Document43 pagesHuman Development Days 1-4Kriztine Mae GonzalesNo ratings yet
- Lab Exercise19 Reproductive System PDFDocument3 pagesLab Exercise19 Reproductive System PDFcsolutionNo ratings yet
- Asc101 Lab Exercise No. 3Document10 pagesAsc101 Lab Exercise No. 3MELITO JR. CATAYLONo ratings yet
- Developmental Biology: GametogenesisDocument21 pagesDevelopmental Biology: GametogenesisTapan Kumar PalNo ratings yet
- Marine Biological Laboratory Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Biological BulletinDocument21 pagesMarine Biological Laboratory Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Biological BulletinRayara Matos Fontineles GuedesNo ratings yet
- 6i. Male Reproduction - OnPRC Module 2-Student HandoutDocument12 pages6i. Male Reproduction - OnPRC Module 2-Student HandoutAkash ShawNo ratings yet
- Journal of Morphology - 2020 - Starck - Morphology of The Avian Yolk SacDocument14 pagesJournal of Morphology - 2020 - Starck - Morphology of The Avian Yolk SacMashai LesenyehoNo ratings yet
- Cortical Granule FunctionDocument17 pagesCortical Granule FunctionEric PhillipsNo ratings yet
- Science Aax7890 PDFDocument9 pagesScience Aax7890 PDFAndoingNo ratings yet
- Acrosome ReactionDocument4 pagesAcrosome ReactionKuldip DwivediNo ratings yet
- 1999 Ency HessDocument8 pages1999 Ency HessKishan GaurNo ratings yet
- Role of Glycolysis and Respiration in Sperm Metabolism and MotilityDocument60 pagesRole of Glycolysis and Respiration in Sperm Metabolism and MotilitywalczakcNo ratings yet
- Fertilization Overview: Sperm PenetrationDocument13 pagesFertilization Overview: Sperm PenetrationAmira FahruddinNo ratings yet
- Biology - Human ReproductionDocument21 pagesBiology - Human Reproductionsgw67No ratings yet
- LAB EXERCISE 2 - OogenesisDocument4 pagesLAB EXERCISE 2 - OogenesisGerald Angelo DeguinioNo ratings yet
- Biology of Polyspermy in IVF and Its Clinical IndicationDocument6 pagesBiology of Polyspermy in IVF and Its Clinical IndicationMD LarasatiNo ratings yet
- Transcriptional Profiling and Physiological Roles of Aedes Aegypti Spermathecal-Related GenesDocument18 pagesTranscriptional Profiling and Physiological Roles of Aedes Aegypti Spermathecal-Related GenesSara VillaNo ratings yet
- tmpD7B9 TMPDocument12 pagestmpD7B9 TMPFrontiersNo ratings yet
- (Unit 4) Human GametesDocument8 pages(Unit 4) Human GametesHanif AlfarizkiNo ratings yet
- Bao Quan Mo Tinh HoanDocument21 pagesBao Quan Mo Tinh Hoanhhhhhhhhhh0000No ratings yet
- Lab Report 4 GametogenesisDocument13 pagesLab Report 4 GametogenesisWnz NaiveNo ratings yet
- Investigating Reproducibility and Tracking ProvenaDocument15 pagesInvestigating Reproducibility and Tracking ProvenaAhmad SolihinNo ratings yet
- Best Practices For Variant Calling in Clinical Sequencing: Review Open AccessDocument13 pagesBest Practices For Variant Calling in Clinical Sequencing: Review Open AccessAhmad SolihinNo ratings yet
- Assessment of The Genomic Variation in A Cattle Population by Re-Sequencing of Key Animals at Low To Medium CoverageDocument9 pagesAssessment of The Genomic Variation in A Cattle Population by Re-Sequencing of Key Animals at Low To Medium CoverageAhmad SolihinNo ratings yet
- ID Pengaruh Propylthiuouracil Terhadap KadaDocument8 pagesID Pengaruh Propylthiuouracil Terhadap KadaAhmad SolihinNo ratings yet
- 1 s2.0 S0300483X20301670 MainDocument12 pages1 s2.0 S0300483X20301670 MainAhmad SolihinNo ratings yet
- Steroids: SciencedirectDocument11 pagesSteroids: SciencedirectAhmad SolihinNo ratings yet
- T-ARMS PCR Genotyping of SNP rs445709131 Using Thermostable Strand Displacement PolymeraseDocument5 pagesT-ARMS PCR Genotyping of SNP rs445709131 Using Thermostable Strand Displacement PolymeraseAhmad SolihinNo ratings yet
- Environmental Research: Yingying Yang, Luqing Pan, Yueyao Zhou, Ruiyi Xu, Dongyu LiDocument12 pagesEnvironmental Research: Yingying Yang, Luqing Pan, Yueyao Zhou, Ruiyi Xu, Dongyu LiAhmad SolihinNo ratings yet
- Guide To The Care and Use of Experimental Animals, Volume 2 (1984) Ix. DogsDocument14 pagesGuide To The Care and Use of Experimental Animals, Volume 2 (1984) Ix. DogsAhmad SolihinNo ratings yet
- Food and Chemical ToxicologyDocument10 pagesFood and Chemical ToxicologyAhmad SolihinNo ratings yet
- Testicular Biopsy: Clinical Practice and Interpretation: ReviewDocument6 pagesTesticular Biopsy: Clinical Practice and Interpretation: ReviewAhmad SolihinNo ratings yet
- The Maturation of Murine Spermatozoa Membranes Within The Epididymis A Computational Biology PerspectiveDocument11 pagesThe Maturation of Murine Spermatozoa Membranes Within The Epididymis A Computational Biology PerspectiveAhmad SolihinNo ratings yet
- Sperm Proteome Maturation in The Mouse EpididymisDocument23 pagesSperm Proteome Maturation in The Mouse EpididymisAhmad SolihinNo ratings yet
- Improving Bioscience Research Reporting: The ARRIVE Guidelines For Reporting Animal ResearchDocument5 pagesImproving Bioscience Research Reporting: The ARRIVE Guidelines For Reporting Animal ResearchAhmad SolihinNo ratings yet
- Molecular Biology of Spermatogenesis: Novel Targets of Apparently Idiopathic Male InfertilityDocument13 pagesMolecular Biology of Spermatogenesis: Novel Targets of Apparently Idiopathic Male InfertilityAhmad SolihinNo ratings yet
- Atlas Anatomi Dan Histologi Binatang Part 1Document15 pagesAtlas Anatomi Dan Histologi Binatang Part 1Ahmad SolihinNo ratings yet
- Annals Oi Anatomy - : Apocrine Secretion - Fact or Artifact?Document12 pagesAnnals Oi Anatomy - : Apocrine Secretion - Fact or Artifact?Ahmad SolihinNo ratings yet
- Pi Is 0021925820827409Document8 pagesPi Is 0021925820827409Ahmad SolihinNo ratings yet
- Vmi2011 757194Document13 pagesVmi2011 757194Ahmad SolihinNo ratings yet
- Practica Reproducitve Systme and PresentDocument10 pagesPractica Reproducitve Systme and PresentLauris CassoNo ratings yet
- PennnyssDocument33 pagesPennnyssscribdmefaischierNo ratings yet
- Male Reproductive SystemDocument15 pagesMale Reproductive SystemCharlz ZipaganNo ratings yet
- Physiologi of Reproductive System With Voice - CompressedDocument16 pagesPhysiologi of Reproductive System With Voice - CompressedMsatriaNo ratings yet
- S10 Q3 WEEK1 Reproductive System LECTUREDocument35 pagesS10 Q3 WEEK1 Reproductive System LECTUREREGLOS, Marie Nhelle K.No ratings yet
- Phimosis Cure TestimonyDocument3 pagesPhimosis Cure TestimonyEnrique RojasNo ratings yet
- Seminal Fluid AnalysisDocument11 pagesSeminal Fluid AnalysisKenneth Jake BatiduanNo ratings yet
- ARTIKEL ILMIAH (061611535039 - Novi Fitria Hanifah)Document10 pagesARTIKEL ILMIAH (061611535039 - Novi Fitria Hanifah)novi fitriaNo ratings yet
- Third Assignment (Forensic)Document2 pagesThird Assignment (Forensic)Dean Mark AnacioNo ratings yet
- Anatomy of The Male Reproductive SystemDocument70 pagesAnatomy of The Male Reproductive SystemDaniel HikaNo ratings yet
- 02 Overview of The Male and Female Reproductive SystemsDocument30 pages02 Overview of The Male and Female Reproductive SystemsOsman Saidu SesayNo ratings yet
- Ovozoa Vol. 7, No. 2, Oktober 2018 ISSN: 2302-6464Document6 pagesOvozoa Vol. 7, No. 2, Oktober 2018 ISSN: 2302-6464Miftah LailiNo ratings yet
- O'Hara & O'Hara - The Effect of Male Circumcision On The Sexual Enjoyment of The Female Partner PDFDocument6 pagesO'Hara & O'Hara - The Effect of Male Circumcision On The Sexual Enjoyment of The Female Partner PDFOmar MohammedNo ratings yet
- Anatomy For Beginners - Reproduction WorksheetDocument4 pagesAnatomy For Beginners - Reproduction WorksheetJames Dauray100% (1)
- Pourusha VastiDocument54 pagesPourusha Vastiksr prasadNo ratings yet
- The PenisDocument6 pagesThe Penisntambik21No ratings yet
- Pengaruh Suhu Dan Lama Thawing Di Dataran Rendah Terhadap Kualitas Semen Beku Sapi BrahmanDocument7 pagesPengaruh Suhu Dan Lama Thawing Di Dataran Rendah Terhadap Kualitas Semen Beku Sapi BrahmanEnikaaNo ratings yet
- Kelly9e Ch03 Male Sexual-Anatomy and PhysiologyDocument19 pagesKelly9e Ch03 Male Sexual-Anatomy and Physiologysauloalberto6254No ratings yet
- Histology Male ReproductiveDocument31 pagesHistology Male Reproductivekhiks34No ratings yet
- Be My Temptation (COMPLETED) - Shaleh - Wattpad - WattpadDocument256 pagesBe My Temptation (COMPLETED) - Shaleh - Wattpad - WattpadWeef hoofNo ratings yet
- 2.1.1 Reproductive Systems Teacher Answer SheetDocument3 pages2.1.1 Reproductive Systems Teacher Answer SheetFerdi Hasan100% (1)
- SpermDocument1 pageSpermEve Karen PobleteNo ratings yet
- Abordul Laparoscopic Pentru Testicule Nepalpabile Aspecte Tehnice Si RezultateDocument6 pagesAbordul Laparoscopic Pentru Testicule Nepalpabile Aspecte Tehnice Si RezultateAlexandra NechiforNo ratings yet
- Sperm Maturation in The Domestic CatDocument11 pagesSperm Maturation in The Domestic CatGeovanni LunaNo ratings yet
- Histologi Reproduksi PriaDocument45 pagesHistologi Reproduksi Priablack junoNo ratings yet
- Maximum Sex! by Editors of MaximDocument200 pagesMaximum Sex! by Editors of MaximLoto760% (3)
- Anatomy of The Male Reproductive SystemDocument24 pagesAnatomy of The Male Reproductive SystemAbbasi KahumaNo ratings yet
- Male Reproductive SystemDocument10 pagesMale Reproductive SystemTracy zorcaNo ratings yet
- Paraphimosis (Ingles)Document1 pageParaphimosis (Ingles)Joaquín SosaNo ratings yet
- Developmental Process of SpermatogenesisDocument4 pagesDevelopmental Process of Spermatogenesispricilia ardianiNo ratings yet