Download as pdf or txt
You might also like
- 2013 A Level H2 Chem Paper 2 Suggested Solutions and Comments PDFDocument8 pages2013 A Level H2 Chem Paper 2 Suggested Solutions and Comments PDFImagreenbucklegirl SGNo ratings yet
- Manufacture of Nitric Acid by The Oxidation of Ammonia'Document6 pagesManufacture of Nitric Acid by The Oxidation of Ammonia'Aqib SiddiqueNo ratings yet
- Culture of Marine PolychaeteDocument7 pagesCulture of Marine PolychaeteyaumilakbarNo ratings yet
- Mussel Adhesion - Essential Footwork: J. Herbert WaiteDocument14 pagesMussel Adhesion - Essential Footwork: J. Herbert WaiteKhalid AhmedNo ratings yet
- Parasite 199065 S 1 P 37Document6 pagesParasite 199065 S 1 P 37Bala Murali KrishnanNo ratings yet
- 1 s2.0 S1319562X17301079 MainDocument8 pages1 s2.0 S1319562X17301079 MainLidwina Faraline TriprisilaNo ratings yet
- Merizocotyle Australensis Himantura FaiDocument12 pagesMerizocotyle Australensis Himantura FaiAlexander MoralesNo ratings yet
- Locations of Gut Associated Lymphoid Tissue in The 3 Month Old Chicken A Review PDFDocument9 pagesLocations of Gut Associated Lymphoid Tissue in The 3 Month Old Chicken A Review PDFDulce AmorNo ratings yet
- Porifera: Sally P. Leys and Nathan FarrarDocument9 pagesPorifera: Sally P. Leys and Nathan FarrarFaradilla Aqidatul IzzahNo ratings yet
- Bacteria TermitasDocument8 pagesBacteria TermitasRashaverak RomNo ratings yet
- Chloroplast SymbiosisDocument10 pagesChloroplast SymbiosisMaria Quesada BrunaNo ratings yet
- 5 PoriferaDocument14 pages5 PoriferaGonzalo Oyanguren AparicioNo ratings yet
- Kalinina Protistology 12-1Document9 pagesKalinina Protistology 12-1jesus javier hoyos acuñaNo ratings yet
- Review Water Relations of Tetrapod Integument: Harvey B. LillywhiteDocument25 pagesReview Water Relations of Tetrapod Integument: Harvey B. LillywhiteDesy NurprasetiyoNo ratings yet
- Kershaw Et Al., 2018 - Understanding Palaeozoic Stromatoporoid GrowthDocument24 pagesKershaw Et Al., 2018 - Understanding Palaeozoic Stromatoporoid Growthjoshuaseo0123No ratings yet
- Coprophilous FungiDocument4 pagesCoprophilous FungiIo DobriNo ratings yet
- PhaeophyceaeDocument4 pagesPhaeophyceaeAdi WeerasingheNo ratings yet
- Examples of CnidariaDocument5 pagesExamples of CnidariaPeh Jia huiNo ratings yet
- Capture, Swallowing, and Egestion of Microplastics by A PlanktivorousDocument8 pagesCapture, Swallowing, and Egestion of Microplastics by A Planktivorousmegha deyNo ratings yet
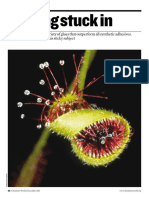
- Bioadhesives - Getting Stuck in - tcm18-210693Document4 pagesBioadhesives - Getting Stuck in - tcm18-210693Hector ReNo ratings yet
- Sindy BeckerDocument11 pagesSindy BeckerHafidz GalantNo ratings yet
- Research Journal of Pharmaceutical, Biological and Chemical SciencesDocument10 pagesResearch Journal of Pharmaceutical, Biological and Chemical SciencesVenu Gopal NNo ratings yet
- CoelenterataDocument27 pagesCoelenterataMia LuvIcecream Part IINo ratings yet
- Entamoeba 4Document3 pagesEntamoeba 4Shyenette SarmientoNo ratings yet
- Lab 5Document3 pagesLab 5alyssa cambaNo ratings yet
- Bio 320 Practical 7Document12 pagesBio 320 Practical 7MuhammadAsyraf67% (3)
- Vesicular, Ulcerative, and Necrotic Dermatitis of ReptilesDocument21 pagesVesicular, Ulcerative, and Necrotic Dermatitis of ReptilesAlice RmaNo ratings yet
- CHDocument2 pagesCHapi-444439435No ratings yet
- Laboratory 3 Cnidaria and CtenophoraDocument13 pagesLaboratory 3 Cnidaria and Ctenophorapokesurfer100% (1)
- Lab 9Document3 pagesLab 9alyssa cambaNo ratings yet
- Chapter5 DiatomsInRivers Burliga Kociolek 2016Document36 pagesChapter5 DiatomsInRivers Burliga Kociolek 2016Thomas ChurchNo ratings yet
- Artículo JellyfishDocument15 pagesArtículo JellyfishNathalie Montserrat Guadarrama SantiagoNo ratings yet
- Ticks and Tick-Borne DiseasesDocument8 pagesTicks and Tick-Borne DiseasesLørïïs SïerraNo ratings yet
- Chapter 17. Bothriocephalus Acheilognathi Yamaguti, 1934: December 2012Document16 pagesChapter 17. Bothriocephalus Acheilognathi Yamaguti, 1934: December 2012Igor YuskivNo ratings yet
- Art3Anatomia ComparadaDocument13 pagesArt3Anatomia ComparadaOscar Hernandez HenaoNo ratings yet
- Saborowskietal 2022Document10 pagesSaborowskietal 2022keltony aquinoNo ratings yet
- Fossil Invertebrates: ArthropodsDocument8 pagesFossil Invertebrates: ArthropodsMuhammad Alfid AlquddusNo ratings yet
- Zoology 1Document4 pagesZoology 1rotoho1793No ratings yet
- Cestodes and TrematodesDocument32 pagesCestodes and TrematodesDenise Sta. AnaNo ratings yet
- Lecture 15 Notes 2013Document5 pagesLecture 15 Notes 2013Richard HampsonNo ratings yet
- Parra2009 PDFDocument9 pagesParra2009 PDFJorge Luis SantamariaNo ratings yet
- Fate Microplastic DemospongeDocument8 pagesFate Microplastic Demospongexnein11No ratings yet
- 6b. Nematode AnatDocument5 pages6b. Nematode Anatmariamagdalena23No ratings yet
- Sheep PaperDocument10 pagesSheep Paperbilal1311511382No ratings yet
- MolluscsDocument26 pagesMolluscslau.wejarNo ratings yet
- The Filter-House of The Larvacean Oikopleura DioicDocument15 pagesThe Filter-House of The Larvacean Oikopleura Dioicromaehab201912No ratings yet
- U1-Volland Et Al 2022 (A Centimeter-Long Bacterium With DNA Compartmentalized in Membrane-Bound Organelles)Document8 pagesU1-Volland Et Al 2022 (A Centimeter-Long Bacterium With DNA Compartmentalized in Membrane-Bound Organelles)LaraKielmasNo ratings yet
- Parasitology Parasitic Helminthes PDFDocument14 pagesParasitology Parasitic Helminthes PDFJuliana MendozaNo ratings yet
- Tapeworms: Jay Georgi, D.V.M., Ph.D.Document21 pagesTapeworms: Jay Georgi, D.V.M., Ph.D.Ana VargasNo ratings yet
- Filum Rotifera-Brusca Et Al., 2019Document24 pagesFilum Rotifera-Brusca Et Al., 2019Madison1920No ratings yet
- 1 ForaminiferaDocument51 pages1 ForaminiferaManoj KitchappanNo ratings yet
- Further Twists in Gastropod Shell EvolutionDocument4 pagesFurther Twists in Gastropod Shell EvolutionLeoregine RodriguezNo ratings yet
- Lom2003 MicrosporidiaFishPhylogenDocument21 pagesLom2003 MicrosporidiaFishPhylogenJorge Manuel Cardenas CallirgosNo ratings yet
- Richman Jackson 1992 A Review of The Ethology of Jumping SpidersDocument5 pagesRichman Jackson 1992 A Review of The Ethology of Jumping SpidersprasanNo ratings yet
- Enperiment For Spotting-3Document18 pagesEnperiment For Spotting-3Debayan Bhattacharyya class:- 11-ANo ratings yet
- PilotwhaleDocument5 pagesPilotwhalejuliette Soubirou-blotNo ratings yet
- Expert Opinion On Drug Delivery Volume 3 Issue 6 2006 (Doi 10.1517/17425247.3.6.727) Benson, Heather AE - Transfersomes For Transdermal Drug DeliveryDocument11 pagesExpert Opinion On Drug Delivery Volume 3 Issue 6 2006 (Doi 10.1517/17425247.3.6.727) Benson, Heather AE - Transfersomes For Transdermal Drug DeliveryprinceamitNo ratings yet
- MacroalgasDocument13 pagesMacroalgas0244761No ratings yet
- Toe Pad Litoria ArticleDocument9 pagesToe Pad Litoria Articlegummy25No ratings yet
- Crop Physiology Exercise-2Document7 pagesCrop Physiology Exercise-2Ravikant MishraNo ratings yet
- SynapsidsDocument7 pagesSynapsidsapi-407347220No ratings yet
- New Distribution of Rhinoceros Beetle Xylotrupes TDocument4 pagesNew Distribution of Rhinoceros Beetle Xylotrupes TMuhammad HazmiNo ratings yet
- JFishBiol 2016 EAFCDocument16 pagesJFishBiol 2016 EAFCMuhammad HazmiNo ratings yet
- 1 SMDocument6 pages1 SMMuhammad HazmiNo ratings yet
- Development of A Multilocus-Based Approach For Sponge (Phylum Porifera) Identification: Refinement and LimitationsDocument14 pagesDevelopment of A Multilocus-Based Approach For Sponge (Phylum Porifera) Identification: Refinement and LimitationsMuhammad HazmiNo ratings yet
- Jurnal SishewDocument13 pagesJurnal SishewMuhammad HazmiNo ratings yet
- Jurnal EnglishDocument6 pagesJurnal EnglishMuhammad HazmiNo ratings yet
- Groups, Noble Gases, Metals, Non-Metals and Metalloids. Transition ElementDocument3 pagesGroups, Noble Gases, Metals, Non-Metals and Metalloids. Transition ElementEman Tan DuNo ratings yet
- (Xii) Alcohol, Phenol, EtherDocument10 pages(Xii) Alcohol, Phenol, EtherAbhishek SharmaNo ratings yet
- Chem P N D Numero 3Document3 pagesChem P N D Numero 3BrittanyNo ratings yet
- iGCSE - Chem - Worksheet 20 - RatesDocument3 pagesiGCSE - Chem - Worksheet 20 - Rateskashif mohammedNo ratings yet
- Food Chemistry A Laboratory Manual 2nd EditionDocument345 pagesFood Chemistry A Laboratory Manual 2nd EditionJawher Bouadila100% (2)
- Biocatalysis NtsDocument2 pagesBiocatalysis NtsBhadra GNo ratings yet
- Kendriya Vidyalaya VirudhunagarDocument18 pagesKendriya Vidyalaya Virudhunagarthejahsree rajaridhuNo ratings yet
- UNIFAC - Parámetros de Volumen y AreaDocument4 pagesUNIFAC - Parámetros de Volumen y Areaisabella MartinezNo ratings yet
- Control of Tacky Deposits On Paper Machines - A Review PDFDocument18 pagesControl of Tacky Deposits On Paper Machines - A Review PDFBiantoroKunartoNo ratings yet
- Jee Chem 1 Eng 26 03Document4 pagesJee Chem 1 Eng 26 03vikasgnsharma100% (1)
- IMORTANT QuestionsDocument5 pagesIMORTANT QuestionsKaran KatamNo ratings yet
- Polarography Introduction and History:: 1922 by The Czech Chemist Jaroslav Heyrovský (1890-1967)Document11 pagesPolarography Introduction and History:: 1922 by The Czech Chemist Jaroslav Heyrovský (1890-1967)Hira AnwarNo ratings yet
- Corbett Quevar Report Oct 2009 FNLDocument25 pagesCorbett Quevar Report Oct 2009 FNLWilliam Arenas BustillosNo ratings yet
- Authors Full Name Abubakar Saleem Muhamm PDFDocument105 pagesAuthors Full Name Abubakar Saleem Muhamm PDFpradyum soniNo ratings yet
- Densities of Common SolidsDocument9 pagesDensities of Common SolidsdeshpravinNo ratings yet
- TUT 3 GC SolutionsDocument4 pagesTUT 3 GC SolutionsLondeka ButheleziNo ratings yet
- Wo 2010038953Document18 pagesWo 2010038953Ranjana MadabushiNo ratings yet
- 7 - Topic 7 - Rate of Chemical ReactionsDocument38 pages7 - Topic 7 - Rate of Chemical Reactionsabud.shawa1No ratings yet
- (Nanostructure Science and Technology) Guozhen Shen, Yu-Lun Chueh - Nanowire Electronics-Springer Singapore (2019)Document396 pages(Nanostructure Science and Technology) Guozhen Shen, Yu-Lun Chueh - Nanowire Electronics-Springer Singapore (2019)Magzoub NourNo ratings yet
- Novel Approach of silica-PVA Hybrid Aerogel Synthesis by Simultaneous Sol-Gel Process and Phase SeparationDocument9 pagesNovel Approach of silica-PVA Hybrid Aerogel Synthesis by Simultaneous Sol-Gel Process and Phase SeparationVeronica HernandezNo ratings yet
- Electrophoretic Deposition of MaterialsDocument27 pagesElectrophoretic Deposition of Materialshichiku4uNo ratings yet
- High Voltage Insulators PDFDocument8 pagesHigh Voltage Insulators PDFKhaled IbrahimNo ratings yet
- TCP 2 (Dyeing&Amp Printing)Document158 pagesTCP 2 (Dyeing&Amp Printing)Kamini PatilNo ratings yet
- Physical Properties of Nylon 6-10Document3 pagesPhysical Properties of Nylon 6-10sharmi2011No ratings yet
- Prevent CrackingDocument2 pagesPrevent CrackingRafeek ShaikhNo ratings yet
- 1 Catalyst FundamentalsDocument17 pages1 Catalyst FundamentalsSam AnuNo ratings yet
- (CSEC Chemistry) Section A Notes and ESQsDocument191 pages(CSEC Chemistry) Section A Notes and ESQsNathaniel WhyteNo ratings yet
- Group 3 Postlab LipidsDocument2 pagesGroup 3 Postlab LipidsSVPSNo ratings yet