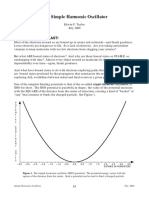
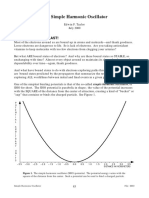
Download as docx, pdf, or txt
You might also like
- French Raven (1959) PDFDocument11 pagesFrench Raven (1959) PDFTuấn PhápNo ratings yet
- CRC Quiz - Theories of CounsellingDocument3 pagesCRC Quiz - Theories of CounsellingIlanna100% (4)
- 1935 - Two Types of Conditioned Reflex and A Pseudo TypeDocument9 pages1935 - Two Types of Conditioned Reflex and A Pseudo TypemchlspNo ratings yet
- Skinner, B. F. (1933) - The Abolishment of A Discrimination PDFDocument4 pagesSkinner, B. F. (1933) - The Abolishment of A Discrimination PDFJota S. FernandesNo ratings yet
- Skinner, B. F. (1934) .The Extinction of Chained Reflexes PDFDocument4 pagesSkinner, B. F. (1934) .The Extinction of Chained Reflexes PDFJota S. FernandesNo ratings yet
- Razran - The Law of Effect or The Law of Qualitative ConditioningDocument19 pagesRazran - The Law of Effect or The Law of Qualitative ConditioningNatalia FfGg100% (1)
- The Associative Structure of Instrumental ConditioningDocument26 pagesThe Associative Structure of Instrumental ConditioningKM SajathNo ratings yet
- Psychological Perspectives On Elt (1) - BehaviourismDocument22 pagesPsychological Perspectives On Elt (1) - BehaviourismALEENA ROSE VARGHESENo ratings yet
- Teorie Relativistiche: Dispensa 1 - Four Steps For Deriving Lorentz' TransformationsDocument9 pagesTeorie Relativistiche: Dispensa 1 - Four Steps For Deriving Lorentz' TransformationsGiacomina SgròNo ratings yet
- TWO TYPES OF CONDITIONED REFLEX, A REPLY TO KONORSKI AND MILLER B.F. - Skinner (1937)Document13 pagesTWO TYPES OF CONDITIONED REFLEX, A REPLY TO KONORSKI AND MILLER B.F. - Skinner (1937)LeoprotNo ratings yet
- NLD UpgradeDocument6 pagesNLD UpgradeAtharv TambadeNo ratings yet
- Yasui 1997Document9 pagesYasui 1997Mohammad AhedNo ratings yet
- Pendulo 1Document10 pagesPendulo 1Geovana BetteroNo ratings yet
- Lec 40Document20 pagesLec 40Manash SarmahNo ratings yet
- Article - 2000 - Non-Smooth Mechanical SystemsDocument8 pagesArticle - 2000 - Non-Smooth Mechanical SystemsmoslemNo ratings yet
- Skinner (1938) - The Behavior of OrganismsDocument486 pagesSkinner (1938) - The Behavior of OrganismsMatheus Jose Schnaider0% (1)
- Chemical Reaction Engineering Prof. Jayant Modak Department of Chemical Engineerin.g Indian Institute of Science, BangaloreDocument17 pagesChemical Reaction Engineering Prof. Jayant Modak Department of Chemical Engineerin.g Indian Institute of Science, Bangaloreary.engenharia1244No ratings yet
- Decay Constant: Itisarate Of1 Order Reaction, It's Unit Is Second Its Value Doesn't Depend On TemperatureDocument5 pagesDecay Constant: Itisarate Of1 Order Reaction, It's Unit Is Second Its Value Doesn't Depend On TemperaturePranabes BhattacharyyaNo ratings yet
- Chapter 6 - 542Document26 pagesChapter 6 - 542Nada KhanNo ratings yet
- BK Pandey 01Document70 pagesBK Pandey 01Anurag SrivastavaNo ratings yet
- Subject ChemistryDocument13 pagesSubject Chemistrysatheeshpharma6No ratings yet
- Chemical Reviews: The Hard Acids Bases (HSAB) Principle and Organic ChemistryDocument20 pagesChemical Reviews: The Hard Acids Bases (HSAB) Principle and Organic ChemistryPinottoNo ratings yet
- Second and First Order Phase Transition Analogy in The Operation of An Organic Dye LaserDocument3 pagesSecond and First Order Phase Transition Analogy in The Operation of An Organic Dye LasercesarNo ratings yet
- Flip FlopDocument25 pagesFlip FlopAjay SahreNo ratings yet
- Relaxation Oscillation Dynamics in Semiconductor Diode Lasers With Optical FeedbackDocument3 pagesRelaxation Oscillation Dynamics in Semiconductor Diode Lasers With Optical FeedbackdabalejoNo ratings yet
- Lecture 11 ADocument7 pagesLecture 11 ARainesius DohlingNo ratings yet
- Research Article: Global Stability of Two-Group Epidemic Models With Distributed Delays and Random PerturbationDocument13 pagesResearch Article: Global Stability of Two-Group Epidemic Models With Distributed Delays and Random PerturbationAndre PNo ratings yet
- Pass Along The Chain. Since Neighbouring Spins Have Become MisalignedDocument1 pagePass Along The Chain. Since Neighbouring Spins Have Become MisalignedKetanNo ratings yet
- Joel H. Villegas: For Your Last Assignment, Please Provide The Following Info. About WaterDocument3 pagesJoel H. Villegas: For Your Last Assignment, Please Provide The Following Info. About WaterJOEL VILLEGASNo ratings yet
- Lec 15Document16 pagesLec 15ary.engenharia1244No ratings yet
- 7.02 SN2 Reaction Mechanism Energy Diagram and StereochemistryDocument3 pages7.02 SN2 Reaction Mechanism Energy Diagram and StereochemistryElizabeth ElsaNo ratings yet
- 05.1 Orbits in Spherical PotentialsDocument26 pages05.1 Orbits in Spherical PotentialsSam PicassoNo ratings yet
- An Approach To The Determination of Reaction in Statically Determinate BeamsDocument4 pagesAn Approach To The Determination of Reaction in Statically Determinate BeamsAbba-Gana MohammedNo ratings yet
- CH 4 Thermo May 2010Document37 pagesCH 4 Thermo May 2010Jayden WangNo ratings yet
- Experiment 2: Kinetics of A Reversible, First-Order, Consecutive ReactionDocument8 pagesExperiment 2: Kinetics of A Reversible, First-Order, Consecutive ReactionVinicius GuimarãesNo ratings yet
- Pericyclic Reactions: Carey & Sundberg: Part A Chapter 11 Carey & Sundberg: Part B Chapter 6 Fleming, Chapter 4Document55 pagesPericyclic Reactions: Carey & Sundberg: Part A Chapter 11 Carey & Sundberg: Part B Chapter 6 Fleming, Chapter 4shoeb321No ratings yet
- Redox ReactionsDocument8 pagesRedox ReactionsHassanNo ratings yet
- Second Order Reactions KineticsDocument8 pagesSecond Order Reactions KineticsPrasad AlexNo ratings yet
- Factors Affecting Rate of Nucleophilic Substitution Reactions Designing A "Good" Nucleophilic SubstitutionDocument9 pagesFactors Affecting Rate of Nucleophilic Substitution Reactions Designing A "Good" Nucleophilic SubstitutionchuasioklengNo ratings yet
- Nucleophilic Substitution Reactions PDFDocument13 pagesNucleophilic Substitution Reactions PDFBhushan Dravyakar100% (6)
- Fluorescence and PhosphorescenceDocument9 pagesFluorescence and PhosphorescenceAliya khalilNo ratings yet
- Phase EquilibriaDocument18 pagesPhase EquilibriaA S SahashransuNo ratings yet
- EconometricsDocument22 pagesEconometricsXheeraz Xaleem NahiyoonNo ratings yet
- 14-2 Effects of Electron-Electron Repulsion: 14-3 Exclusion Principle and Exchange InteractionDocument4 pages14-2 Effects of Electron-Electron Repulsion: 14-3 Exclusion Principle and Exchange InteractionFina Aulia RitongaNo ratings yet
- Unit 5 Fluorimetry and Phosphorimetry-1Document23 pagesUnit 5 Fluorimetry and Phosphorimetry-1Saif YounusNo ratings yet
- Reaksi Senyawa Kompleks PoncoDocument13 pagesReaksi Senyawa Kompleks PoncoSeftiani YusniaNo ratings yet
- Raman Spectral NotesDocument4 pagesRaman Spectral NotesswethasNo ratings yet
- Haloalkanes and Alcohols Nucleophilic Substitution and Elimination ReactionsDocument4 pagesHaloalkanes and Alcohols Nucleophilic Substitution and Elimination ReactionsanonymousNo ratings yet
- Synchronous Machine Modeling - Notes2 1.0 Inductances: I I L L L L L L L L L L L L L L L L L LDocument25 pagesSynchronous Machine Modeling - Notes2 1.0 Inductances: I I L L L L L L L L L L L L L L L L L LAlexander MartinNo ratings yet
- I.: J. F. J.: Rheological Properties O F BitumensDocument17 pagesI.: J. F. J.: Rheological Properties O F BitumensoreamigNo ratings yet
- Atomic Term SymbolsDocument13 pagesAtomic Term SymbolsAnish RaoNo ratings yet
- The Simple Harmonic Oscillator: Bound States - at Last!Document6 pagesThe Simple Harmonic Oscillator: Bound States - at Last!abcofzNo ratings yet
- The Simple Harmonic Oscillator: Bound States - at Last!Document6 pagesThe Simple Harmonic Oscillator: Bound States - at Last!Marcos SilvaNo ratings yet
- Exam 1 Chem 102 in ColorDocument6 pagesExam 1 Chem 102 in ColorKevinZamudioNo ratings yet
- ThermodynamicsDocument24 pagesThermodynamicsrejie magnayeNo ratings yet
- 05 Atmospheric DynamicsDocument37 pages05 Atmospheric DynamicszhangyuluNo ratings yet
- CFE - 10 - Chemical Kinetics Part 2Document19 pagesCFE - 10 - Chemical Kinetics Part 2Christian C. SuaseNo ratings yet
- Introduction to Non-Linear Mechanics. (AM-11), Volume 11From EverandIntroduction to Non-Linear Mechanics. (AM-11), Volume 11No ratings yet
- Automorphic Representation of Unitary Groups in Three Variables. (AM-123), Volume 123From EverandAutomorphic Representation of Unitary Groups in Three Variables. (AM-123), Volume 123No ratings yet
- Psychoanalytic Basis of MotivationDocument4 pagesPsychoanalytic Basis of MotivationJoshua TuguinayNo ratings yet
- Theories of Personality NOTESDocument17 pagesTheories of Personality NOTEStaylor alison swiftNo ratings yet
- Psychoanalysis 3Document10 pagesPsychoanalysis 3Dolly RizaldoNo ratings yet
- Reality TherapyDocument1 pageReality TherapyTinoRepaso100% (1)
- Cognitive Behavioral Therapy - 2007 MayDocument43 pagesCognitive Behavioral Therapy - 2007 Mayeryxsp100% (2)
- Prod - Ed FaciDocument19 pagesProd - Ed Facijean gonzaga100% (1)
- TFN ReportDocument2 pagesTFN ReportLOUIE ENRIQUEZNo ratings yet
- HPGD 2103 - Guidance and CounsellingDocument14 pagesHPGD 2103 - Guidance and CounsellingkhugeshNo ratings yet
- Cognitive Theory and Associated TherapiesDocument3 pagesCognitive Theory and Associated TherapiesEfren RJ PongaseNo ratings yet
- BK SkinnerDocument5 pagesBK SkinnerJananee RajagopalanNo ratings yet
- Art Therapy: Venue: Maple Musicology Academy Date: 25/3/15 Time: 9am - 5pmDocument14 pagesArt Therapy: Venue: Maple Musicology Academy Date: 25/3/15 Time: 9am - 5pmKyleMichaelisNo ratings yet
- MA SEM II Positive PsychologyDocument130 pagesMA SEM II Positive PsychologyMitesh HarmalkarNo ratings yet
- Daftar Pustaka - 1Document2 pagesDaftar Pustaka - 1IrmaaNo ratings yet
- Burn Out 1Document12 pagesBurn Out 1Rais IrfanNo ratings yet
- ALS ASSESSMENT FORM 1 and 2Document2 pagesALS ASSESSMENT FORM 1 and 2Joecel Dumagpi-Reble100% (1)
- Leeder 2022 Behaviorism Skinnerand Operant Conditioning Considerationsfor Sport Coaching PracticeDocument8 pagesLeeder 2022 Behaviorism Skinnerand Operant Conditioning Considerationsfor Sport Coaching PracticepandayfaelNo ratings yet
- Theories of Psychological DevelopmentDocument19 pagesTheories of Psychological DevelopmentEliha NarineNo ratings yet
- AP Psychology Learning PPT-1Document42 pagesAP Psychology Learning PPT-1simi duttNo ratings yet
- Future Oriented PsychotherapyDocument10 pagesFuture Oriented PsychotherapyzarrarNo ratings yet
- Module 8 Neo Behaviorism Tolman and BanduraDocument8 pagesModule 8 Neo Behaviorism Tolman and Bandurashiela100% (1)
- HBO Chapter 5 REVIEWERDocument11 pagesHBO Chapter 5 REVIEWERJyan GayNo ratings yet
- Belajar Berdasar Regulasi Diri Dalam PembelajaranDocument14 pagesBelajar Berdasar Regulasi Diri Dalam PembelajaranDati DelianaNo ratings yet
- Learning Theory: Presented by - Dr. Riya Singh Junior Resident 1 Department of PsychiatryDocument34 pagesLearning Theory: Presented by - Dr. Riya Singh Junior Resident 1 Department of PsychiatryRiyaSinghNo ratings yet
- PED3 Lesson 3 Learning TheoriesDocument23 pagesPED3 Lesson 3 Learning TheoriesAxel Rosemarie PlecesNo ratings yet
- Theories of Motivation in Education An Integrative FrameworkDocument35 pagesTheories of Motivation in Education An Integrative FrameworkMarcela Piamonte CruzNo ratings yet
- Ego StatesDocument11 pagesEgo StateslvincentrajkumarNo ratings yet
- HW2 Resubmit MGT4479Document8 pagesHW2 Resubmit MGT4479Giang LêNo ratings yet
- Motivation Theories - Faiza NadeemDocument24 pagesMotivation Theories - Faiza NadeemMuhammad AliNo ratings yet
- CBTDocument21 pagesCBTsavvy_as_98No ratings yet