Download as pdf or txt
You might also like
- General Zoology SyllabusDocument4 pagesGeneral Zoology SyllabusNL R Q DO100% (4)
- K.R. Shivanna, Rajesh Tandon (Auth.) - Reproductive Ecology of Flowering Plants - A Manual-Springer India (2014)Document178 pagesK.R. Shivanna, Rajesh Tandon (Auth.) - Reproductive Ecology of Flowering Plants - A Manual-Springer India (2014)Elisabeth Tölke100% (1)
- Jurnal 2Document8 pagesJurnal 2Dea SyafiraNo ratings yet
- Actin Dynamics in Cell MigrationDocument13 pagesActin Dynamics in Cell Migration王麦No ratings yet
- Miguel-Arribas A, Et Al. NAR 2021Document15 pagesMiguel-Arribas A, Et Al. NAR 2021AntonNo ratings yet
- Loreto, M (2002) - Functional Cooperation Between C-CBL and Src-Like Adaptor Protein 2 in The Negative Regulation of T-Cell Receptor SignalingDocument15 pagesLoreto, M (2002) - Functional Cooperation Between C-CBL and Src-Like Adaptor Protein 2 in The Negative Regulation of T-Cell Receptor SignalingAnadahiNo ratings yet
- 1997-Oncoprotein NetworksDocument14 pages1997-Oncoprotein NetworksyicinenNo ratings yet
- Bba - Molecular Cell Research: SciencedirectDocument9 pagesBba - Molecular Cell Research: SciencedirectHaleem AfsarNo ratings yet
- A Flat BAR Protein Promotes Actin Polymerization Atthe Base of Clathrin-Coated PitsDocument28 pagesA Flat BAR Protein Promotes Actin Polymerization Atthe Base of Clathrin-Coated Pitsm9bpb6frpxNo ratings yet
- Fimmu 03 00130Document18 pagesFimmu 03 00130Isabella1237No ratings yet
- Romero - OctoberDocument10 pagesRomero - OctoberkennethdaveastoNo ratings yet
- Stochastic Dynamics of Membrane Protrusion Mediated by The DOCK180/Rac Pathway in Migrating CellsDocument10 pagesStochastic Dynamics of Membrane Protrusion Mediated by The DOCK180/Rac Pathway in Migrating CellsRose Sky SiriusNo ratings yet
- Nihms 47053Document15 pagesNihms 47053zsyedphd1No ratings yet
- WNT Signaling Multiple PathwaysDocument6 pagesWNT Signaling Multiple PathwaysMayerlis ArrietaNo ratings yet
- 07 01 Protein Conducting Channel Xray BergDocument9 pages07 01 Protein Conducting Channel Xray BergI TECHNo ratings yet
- Hansen Et Al 2015 Hippo SignalingDocument15 pagesHansen Et Al 2015 Hippo SignalingAlabhya DasNo ratings yet
- Hu, 2019 PDFDocument10 pagesHu, 2019 PDFAnisa FirdausNo ratings yet
- 1 s2.0 S2001037022002896 MainDocument13 pages1 s2.0 S2001037022002896 MainEzike Tobe ChriszNo ratings yet
- Erratum TibsDocument1 pageErratum TibsAbdichakour Med HassanNo ratings yet
- PNAS 2008 Manning 9674 9Document6 pagesPNAS 2008 Manning 9674 9labmeeting-developersNo ratings yet
- NIH Public Access: Author ManuscriptDocument35 pagesNIH Public Access: Author ManuscriptPutra UtomoNo ratings yet
- Review Debolle 2004 PdhsDocument5 pagesReview Debolle 2004 PdhsdeniseNo ratings yet
- Kennedy 1989Document4 pagesKennedy 1989Eduardo TarangoNo ratings yet
- Cortactin Binding To F-Actin Revealed by Electron Microscopy and 3D ReconstructionDocument8 pagesCortactin Binding To F-Actin Revealed by Electron Microscopy and 3D ReconstructionLilian Elisa HernándezNo ratings yet
- European Journal of Biochemistry - 2001 - Kalies - Protein Translocation Into The Endoplasmic Reticulum ERDocument5 pagesEuropean Journal of Biochemistry - 2001 - Kalies - Protein Translocation Into The Endoplasmic Reticulum ER12 yashika palNo ratings yet
- Protein Tyrosine Phosphatases As Adhesion ReceptorsDocument8 pagesProtein Tyrosine Phosphatases As Adhesion ReceptorsAlexandra AguilarNo ratings yet
- Transduction Protein Interactions Define Specificity in Signal ProteinDocument22 pagesTransduction Protein Interactions Define Specificity in Signal ProteinnitishpathaniaNo ratings yet
- Nat Immunol 2014navarroDocument8 pagesNat Immunol 2014navarrotri sutrianiNo ratings yet
- Cellular Functions of Long Noncoding Rnas: Run-Wen Yao, Yang Wang and Ling-Ling ChenDocument10 pagesCellular Functions of Long Noncoding Rnas: Run-Wen Yao, Yang Wang and Ling-Ling ChenAmrutha Swathi PriyaNo ratings yet
- Clathrin Now You See Me Now You Don T - 2004 - Developmental CellDocument2 pagesClathrin Now You See Me Now You Don T - 2004 - Developmental Cellhalil sahanNo ratings yet
- Neurocytology I - The Structure of The NeuronDocument46 pagesNeurocytology I - The Structure of The NeuronMaharshi MajmundarNo ratings yet
- Biochimica Et Biophysica Acta: Alexander Plotnikov, Eldar Zehorai, Shiri Procaccia, Rony SegerDocument15 pagesBiochimica Et Biophysica Acta: Alexander Plotnikov, Eldar Zehorai, Shiri Procaccia, Rony Segerlinda ratna watiNo ratings yet
- 2009 - Shaw, FilbertDocument10 pages2009 - Shaw, FilbertAfranio Cogo DestefaniNo ratings yet
- 2020 QC Pathways of TA ProteinsDocument9 pages2020 QC Pathways of TA ProteinsLê Khánh ToànNo ratings yet
- 2004 TSCRT ClonningDocument5 pages2004 TSCRT ClonningArturo RojoNo ratings yet
- The C-Terminus of Prestin Influences Nonlinear Capacitance and Plasma Membrane TargetingDocument10 pagesThe C-Terminus of Prestin Influences Nonlinear Capacitance and Plasma Membrane TargetingAlex WongNo ratings yet
- Claudia Gonzalez Lima 2020Document76 pagesClaudia Gonzalez Lima 2020Ann LourieNo ratings yet
- Genes Dev.-2001-Abraham-2177-96Document21 pagesGenes Dev.-2001-Abraham-2177-96M Faizal RezhaNo ratings yet
- Protein Folding in ERDocument17 pagesProtein Folding in ERAfaq AhmadNo ratings yet
- Blume Jensen2001Document11 pagesBlume Jensen2001yusuf eka maulanaNo ratings yet
- Phosphorylation of P-Glycoprotein by PKA and PKC Modulates Swelling-Activated CL CurrentsDocument9 pagesPhosphorylation of P-Glycoprotein by PKA and PKC Modulates Swelling-Activated CL CurrentsDr-Dalya ShakirNo ratings yet
- T Cell Activation and Signaling: TCR TalinDocument8 pagesT Cell Activation and Signaling: TCR TalinMst Monira KhatonNo ratings yet
- The Nucleus - Express YourselfDocument2 pagesThe Nucleus - Express YourselfbresojiarNo ratings yet
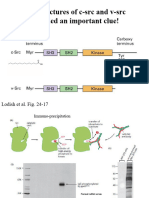
- The Structures of C-SRC and V-SRC Provided An Important Clue!Document33 pagesThe Structures of C-SRC and V-SRC Provided An Important Clue!Souvik DeNo ratings yet
- Tannic Acid Attenuates Vascular Calcification Induced PR - 2021 - Biomedicine PDFDocument12 pagesTannic Acid Attenuates Vascular Calcification Induced PR - 2021 - Biomedicine PDFFernando MaffeisNo ratings yet
- Conserved Microtubule-Actin Interactions in Cell Movement and Morphogenesis 03Document11 pagesConserved Microtubule-Actin Interactions in Cell Movement and Morphogenesis 03Sonia Barbosa CornelioNo ratings yet
- Introduction To The Histology and Cell Biology of The Actin CytoskeletonDocument2 pagesIntroduction To The Histology and Cell Biology of The Actin CytoskeletonpajtgocjNo ratings yet
- Gharbi 2011. DGKZ controlsDAG at ISDocument9 pagesGharbi 2011. DGKZ controlsDAG at ISmcrickboomNo ratings yet
- 1998 Mata2 DegradationDocument11 pages1998 Mata2 DegradationLê Khánh ToànNo ratings yet
- Receptor Interacting Protein Kinase-3Document12 pagesReceptor Interacting Protein Kinase-3cursotplo2021No ratings yet
- Astrina Spectro Masas 2001Document6 pagesAstrina Spectro Masas 2001Nacido para BendcirNo ratings yet
- 560 FullDocument6 pages560 FullAngel AcevedoNo ratings yet
- 2023-NC-Temporal Dynamics of Gene Expression andDocument12 pages2023-NC-Temporal Dynamics of Gene Expression andJay Prakash MauryaNo ratings yet
- Parte 03Document100 pagesParte 03Pâmella PicançoNo ratings yet
- New Insights Into The Regulation and Cellular Functions of The Arp2-3 ComplexDocument6 pagesNew Insights Into The Regulation and Cellular Functions of The Arp2-3 ComplexKarina B Hernandez ANo ratings yet
- Membrane ProteinsDocument3 pagesMembrane ProteinsAfaq AhmadNo ratings yet
- agrin-LRP4-MuSK SignalingDocument12 pagesagrin-LRP4-MuSK SignalingDAVID TERRONESNo ratings yet
- Lee et al 2021Document20 pagesLee et al 2021Arsenii DmitrievNo ratings yet
- Optogenetic Control of The Canonical WNT Signaling Path - 2021 - Journal of MoleDocument12 pagesOptogenetic Control of The Canonical WNT Signaling Path - 2021 - Journal of MolemukundaragaNo ratings yet
- Min 2011Document9 pagesMin 2011Tibor SzénásiNo ratings yet
- 8 Non-Muscle Actin, IFs, Adhesion-2Document35 pages8 Non-Muscle Actin, IFs, Adhesion-2zj5bnxbymzNo ratings yet
- Cell Motility: From Molecules to OrganismsFrom EverandCell Motility: From Molecules to OrganismsAnne RidleyNo ratings yet
- Harvesting of BananaDocument36 pagesHarvesting of Bananaapi-19911503No ratings yet
- Leziunea Acuta de Rinichi (LAR) : Acute Kidney Injury (AKI) Insuficienta Renala Acuta (IRA)Document49 pagesLeziunea Acuta de Rinichi (LAR) : Acute Kidney Injury (AKI) Insuficienta Renala Acuta (IRA)mihai mayorNo ratings yet
- How To Live Longer and Feel Better - Even With CancerDocument21 pagesHow To Live Longer and Feel Better - Even With CancerMaurizio Pieroni0% (1)
- 7.1 Respiratory Process in Energy ProductionDocument13 pages7.1 Respiratory Process in Energy ProductionNor Rafidah Che YusofNo ratings yet
- Descending Tracts: Dr. Niranjan Murthy H L Asst Prof of Physiology SSMC, TumkurDocument23 pagesDescending Tracts: Dr. Niranjan Murthy H L Asst Prof of Physiology SSMC, Tumkurnirilib100% (1)
- Infancy - PPT 1Document26 pagesInfancy - PPT 1Dominic Mathew KNo ratings yet
- Bioluminescence in Flora and Fauna and Its UtilityDocument15 pagesBioluminescence in Flora and Fauna and Its Utilitybabi2798No ratings yet
- Understanding The Human Sexual ResponseDocument12 pagesUnderstanding The Human Sexual ResponseMaan JosueNo ratings yet
- Capitulo 3 Estructura de Las AgallasDocument22 pagesCapitulo 3 Estructura de Las AgallasstraklerNo ratings yet
- Histo SupDocument8 pagesHisto SupAbdoulhaleem MoNo ratings yet
- HIV-1 Proviral DNA Excision Using An Evolved RecombinaseDocument5 pagesHIV-1 Proviral DNA Excision Using An Evolved Recombinaseanon-850655100% (3)
- How Does Miswak Compare To A Toothbrush and Toothpaste in Cleaning and Killing Bacteria?Document1 pageHow Does Miswak Compare To A Toothbrush and Toothpaste in Cleaning and Killing Bacteria?Vishal SoniNo ratings yet
- HematomaDocument2 pagesHematomaLeonel OcanaNo ratings yet
- Econtent PDF EmbryologyGametogenesisDocument9 pagesEcontent PDF EmbryologyGametogenesisVipin BhartiNo ratings yet
- Functional Training TorsoDocument2 pagesFunctional Training TorsoOtrovanjeNo ratings yet
- Sains Bab 1 - Form 2Document7 pagesSains Bab 1 - Form 2THANABALAN A/L MUNUSWAMY MoeNo ratings yet
- Utkalendu Resume-Data ScienceDocument2 pagesUtkalendu Resume-Data Sciencejames neeshamNo ratings yet
- Goprep Talent Search Exam Sample Paper: InstructionsDocument19 pagesGoprep Talent Search Exam Sample Paper: InstructionsBeyond ur imaginationNo ratings yet
- Eshmuno S ResinDocument4 pagesEshmuno S ResinTuyền KimNo ratings yet
- Oxygen Transport JOHN W. BAYNES, MAREK H. DOMINICZAK - Medical Biochemistry-Elsevier Inc. (2019)Document15 pagesOxygen Transport JOHN W. BAYNES, MAREK H. DOMINICZAK - Medical Biochemistry-Elsevier Inc. (2019)Malika MohNo ratings yet
- Plant Genetics and Molecular BiologyDocument30 pagesPlant Genetics and Molecular BiologyCeazar Ian PesimoNo ratings yet
- Histological Characteristics of Human Lingual FrenulumDocument1 pageHistological Characteristics of Human Lingual FrenulumajudNo ratings yet
- EnglishDocument9 pagesEnglishIvy Mae AninonNo ratings yet
- Sorghum Root and Stalk RotsDocument288 pagesSorghum Root and Stalk RotsFindi DiansariNo ratings yet
- Enzyme Purification NotesDocument23 pagesEnzyme Purification NotesMadan Rajan100% (1)
- Power For Every HourDocument10 pagesPower For Every HourmarnesiaNo ratings yet
- Mep 2 Assesemnt CC NewDocument12 pagesMep 2 Assesemnt CC NewPrecious Bardon-MempinNo ratings yet
- Growing Cannabis (The Ultimate Guide) BookDocument69 pagesGrowing Cannabis (The Ultimate Guide) Bookbns.publishing12No ratings yet