Download as pdf or txt
You might also like
- Secrets of Hypnosis Revealed!Document39 pagesSecrets of Hypnosis Revealed!Angel Fulton100% (1)
- Carrom BasicsDocument3 pagesCarrom Basicssurefooted10% (1)
- Leadership ArchitectDocument11 pagesLeadership ArchitectigoeneezmNo ratings yet
- Normal and Pathological Oscillatory Communication in The BrainDocument12 pagesNormal and Pathological Oscillatory Communication in The BrainDiegoNo ratings yet
- Using Goal-Driven Deep Learning Models To Understand The Sensory CortexDocument10 pagesUsing Goal-Driven Deep Learning Models To Understand The Sensory CortexRajsuryan SinghNo ratings yet
- 1 s2.0 S0959438820301355 MainDocument9 pages1 s2.0 S0959438820301355 Maingodray1234aNo ratings yet
- Organic Transistors Mimicking Brain Behavior: K.Sai SameeraDocument6 pagesOrganic Transistors Mimicking Brain Behavior: K.Sai Sameeradba_123No ratings yet
- Computational Neuromodulation Large Scale Neural DynamicsDocument12 pagesComputational Neuromodulation Large Scale Neural DynamicsShayan ShafquatNo ratings yet
- Fields 2015Document12 pagesFields 2015Kornél SzabóNo ratings yet
- Mahvash 2013Document13 pagesMahvash 2013Damian SearaNo ratings yet
- Neuroview: Designing Tools For Assumption-Proof Brain MappingDocument3 pagesNeuroview: Designing Tools For Assumption-Proof Brain Mappingports1111No ratings yet
- CGM 403 NotesDocument6 pagesCGM 403 NotesKrishnaNo ratings yet
- Network Reset: A Simplified Overarching Theory of Locus Coeruleus Noradrenaline FunctionDocument9 pagesNetwork Reset: A Simplified Overarching Theory of Locus Coeruleus Noradrenaline FunctionIsaac Saucedo FloresNo ratings yet
- TMP 1 AEDocument26 pagesTMP 1 AEFrontiersNo ratings yet
- Large-Scale Brain Networks in Cognition: Emerging Methods and PrinciplesDocument14 pagesLarge-Scale Brain Networks in Cognition: Emerging Methods and PrinciplesLuis GómezNo ratings yet
- NPH 011 033403Document28 pagesNPH 011 033403Cristhian RodriguezNo ratings yet
- Pulsed Neural Networks and Their Application: Daniel R. Kunkle Chadd MerriganDocument11 pagesPulsed Neural Networks and Their Application: Daniel R. Kunkle Chadd MerriganinnocentbystanderNo ratings yet
- Ijcnn 2006 246651Document6 pagesIjcnn 2006 246651Triệu HảiNo ratings yet
- Neural Manifold Analysis of Brain Circuit DynamicsDocument24 pagesNeural Manifold Analysis of Brain Circuit DynamicsKarla ValderramaNo ratings yet
- 1 s2.0 S1364661323000645 MainDocument11 pages1 s2.0 S1364661323000645 MainMitia D'AcolNo ratings yet
- Mantini (2007) Electrophysiological Signatures of Resting State Networks in The Human BrainDocument6 pagesMantini (2007) Electrophysiological Signatures of Resting State Networks in The Human BrainJulieht RodriguezNo ratings yet
- Letter: Attention Enhances Synaptic Efficacy and The Signal-To-Noise Ratio in Neural CircuitsDocument8 pagesLetter: Attention Enhances Synaptic Efficacy and The Signal-To-Noise Ratio in Neural Circuitstirin mooreNo ratings yet
- Stimulation of Neuronal Activity With LightDocument29 pagesStimulation of Neuronal Activity With LightThe Panda EntertainerNo ratings yet
- Friston AiDocument15 pagesFriston AiraelpersoneNo ratings yet
- Tripartite Synapse and Active Milieu PDFDocument12 pagesTripartite Synapse and Active Milieu PDFhzol83No ratings yet
- Simulation and Characterization of Neuron Cell Behaviour in FPGADocument8 pagesSimulation and Characterization of Neuron Cell Behaviour in FPGAVaalu SivaNo ratings yet
- Diorio 2003Document7 pagesDiorio 2003diliplokande46No ratings yet
- Gao Et Al. - 2020 - Neuronal Timescales Are Functionally Dynamic and SDocument26 pagesGao Et Al. - 2020 - Neuronal Timescales Are Functionally Dynamic and Srodrigo carlos carrilloNo ratings yet
- End - Sem - For MergeDocument42 pagesEnd - Sem - For Mergeমেঘে ঢাকা তারাNo ratings yet
- 1 s2.0 S0956566323005808 MainDocument11 pages1 s2.0 S0956566323005808 MainLuís CerdeiraNo ratings yet
- Temporal Signal Neuron CircuitDocument14 pagesTemporal Signal Neuron CircuitJacob EveristNo ratings yet
- Infrared Laser Pulses Excite Action Potentials in Primary Cortex Neurons in VitroDocument5 pagesInfrared Laser Pulses Excite Action Potentials in Primary Cortex Neurons in VitroBoros Csanád ÖrsNo ratings yet
- 4 IJAEST Volume No 2 Issue No 1 A Novel Approach To Network Intrusion Detection Using Spiking Neural Networks 036 042Document7 pages4 IJAEST Volume No 2 Issue No 1 A Novel Approach To Network Intrusion Detection Using Spiking Neural Networks 036 042iserpNo ratings yet
- Review: Musical Training As A Framework For Brain Plasticity: Behavior, Function, and StructureDocument17 pagesReview: Musical Training As A Framework For Brain Plasticity: Behavior, Function, and Structureyokai12No ratings yet
- Artificial NNDocument11 pagesArtificial NNkmeghananayakNo ratings yet
- (1977) A Principle of Neural Assoiative MemoryDocument11 pages(1977) A Principle of Neural Assoiative MemorymartindsqNo ratings yet
- Artificial Neural Networks-Based Real-Time Classification of ENG Signals For Implanted Nerve InterfacesDocument16 pagesArtificial Neural Networks-Based Real-Time Classification of ENG Signals For Implanted Nerve InterfacesMark HewittNo ratings yet
- Dynamic BrainDocument35 pagesDynamic BrainchandusgNo ratings yet
- Controllability of Structural Brain Networks: ArticleDocument10 pagesControllability of Structural Brain Networks: ArticleshyamNo ratings yet
- INVITED LECTURE: MIND READING With EEG - Phase Coding/ Decoding - Neurotalk 2010Document35 pagesINVITED LECTURE: MIND READING With EEG - Phase Coding/ Decoding - Neurotalk 2010simon brežanNo ratings yet
- ML cl3 kc1Document11 pagesML cl3 kc1Dr B AnjanadeviNo ratings yet
- Computational Neuroscience 1988-3883Document9 pagesComputational Neuroscience 1988-3883detki007No ratings yet
- Neuromorphic Photonics: Introduction To Spiking and ExcitabilityDocument5 pagesNeuromorphic Photonics: Introduction To Spiking and Excitabilityrama chaudharyNo ratings yet
- Topological Exploration of Artificial Neuronal Network DynamicsDocument20 pagesTopological Exploration of Artificial Neuronal Network DynamicsYishuai ZhangNo ratings yet
- Towards Reverse Engineering Brain Model Abstraction Simulation Framework KrichmarDocument6 pagesTowards Reverse Engineering Brain Model Abstraction Simulation Framework KrichmarAlex WongNo ratings yet
- Invertebrate Nervous Systems: Thomas MathesonDocument6 pagesInvertebrate Nervous Systems: Thomas MathesonMax MoralesNo ratings yet
- Neurobiology RelatedDocument9 pagesNeurobiology RelateddgavrileNo ratings yet
- Untitled - Srep07853Document1 pageUntitled - Srep07853Generation GenerationNo ratings yet
- Buzsaki 2004 Nat NeurosciDocument7 pagesBuzsaki 2004 Nat Neuroscihussin elrashidyNo ratings yet
- Department of Instrumentation EngineeringDocument28 pagesDepartment of Instrumentation EngineeringMahi KatreNo ratings yet
- Three Anti-Correlated Neuronal Networks Managing Brain Activity A ReviewDocument9 pagesThree Anti-Correlated Neuronal Networks Managing Brain Activity A ReviewVioleta-Nella MocanuNo ratings yet
- Innate Synchronous Oscillations in Freely-Organized Small Neuronal CircuitsDocument9 pagesInnate Synchronous Oscillations in Freely-Organized Small Neuronal CircuitsFrontiersNo ratings yet
- Three Anti Correlated Neuronal Networks Managing Brain ActivityDocument8 pagesThree Anti Correlated Neuronal Networks Managing Brain ActivitydgavrileNo ratings yet
- Electrophysiological Brain Connectivity Theory andDocument25 pagesElectrophysiological Brain Connectivity Theory andl.fernandezNo ratings yet
- Sompolinsky - PhysicsToday Statistical Mechanics of Neural NetworksDocument11 pagesSompolinsky - PhysicsToday Statistical Mechanics of Neural Networksanurag sahayNo ratings yet
- SPetkoski Freq2019Document14 pagesSPetkoski Freq2019crunNo ratings yet
- Poster DANUBE 2005 - Working Memory and EEG CoherenceDocument2 pagesPoster DANUBE 2005 - Working Memory and EEG Coherencesimon brežanNo ratings yet
- 2017 - Network NeuroscienceDocument12 pages2017 - Network NeuroscienceMaria Eugenia Amaya ValenciaNo ratings yet
- HCL Shiv VlsiDocument12 pagesHCL Shiv VlsiRavi Teja MutyalaNo ratings yet
- An Evolving Perspective On The Dynamic Brain - Notes From The Brain Conference On Dynamics of The Brain - Temporal Aspects of ComputationDocument22 pagesAn Evolving Perspective On The Dynamic Brain - Notes From The Brain Conference On Dynamics of The Brain - Temporal Aspects of ComputationRajsuryan SinghNo ratings yet
- J Tins 2020 07 001Document13 pagesJ Tins 2020 07 001Iki MikliNo ratings yet
- Coordinated Activity in the Brain: Measurements and Relevance to Brain Function and BehaviorFrom EverandCoordinated Activity in the Brain: Measurements and Relevance to Brain Function and BehaviorJose Luis Perez VelazquezNo ratings yet
- Flexible Neural Electrodes: Revolutionizing Brain-Computer InterfacesFrom EverandFlexible Neural Electrodes: Revolutionizing Brain-Computer InterfacesNo ratings yet
- A Study On Clinical, Radiological and Electrophysiological Profile in Patients Presenting With First Unprovoked SeizureDocument8 pagesA Study On Clinical, Radiological and Electrophysiological Profile in Patients Presenting With First Unprovoked Seizureindex PubNo ratings yet
- Invitation LetterDocument8 pagesInvitation LetterFar SultanNo ratings yet
- CommunicationDocument1 pageCommunicationghrehman_4No ratings yet
- MotivationDocument39 pagesMotivationnavin_tiwari_king100% (1)
- Psychosocial Approaches To Deeply Distur PDFDocument251 pagesPsychosocial Approaches To Deeply Distur PDFFabian R CNo ratings yet
- Aspergers Boarding SchoolDocument4 pagesAspergers Boarding SchoolBrian HaraNo ratings yet
- A Curtis Artifact Sheet Group 4Document3 pagesA Curtis Artifact Sheet Group 4api-363528301No ratings yet
- Computers in Human Behavior: Tracy Packiam Alloway, Ross Geoffrey AllowayDocument7 pagesComputers in Human Behavior: Tracy Packiam Alloway, Ross Geoffrey AllowayKashif KhanNo ratings yet
- Attention Deficit Hyperactivity Disorder: ClassificationDocument14 pagesAttention Deficit Hyperactivity Disorder: ClassificationBharat RasghaniaNo ratings yet
- Tusiani Eng2018Document15 pagesTusiani Eng2018europython100% (1)
- Indian Abacus Racers Question Paper - 4th Level-FreeDocument8 pagesIndian Abacus Racers Question Paper - 4th Level-FreeIndian AbacusNo ratings yet
- Rehabilitation For People With Hemianopia The Next StepDocument7 pagesRehabilitation For People With Hemianopia The Next StepAnonymous k4F8sAs25No ratings yet
- NCP AppendicitisDocument2 pagesNCP AppendicitisArnel Luces Beato100% (1)
- A Model For Art Therapists in Community Based PracticeDocument9 pagesA Model For Art Therapists in Community Based PracticeIrene MaropakisNo ratings yet
- A 62 Years Old Woman With Recurrent Depressive Disorder, Current Episode Moderate, With Somatic SymptomsDocument5 pagesA 62 Years Old Woman With Recurrent Depressive Disorder, Current Episode Moderate, With Somatic SymptomsAkemi Chan CarterNo ratings yet
- J Neurol 2023 Jul 270 (7) 3654-3666Document13 pagesJ Neurol 2023 Jul 270 (7) 3654-3666lucas javier CobraNo ratings yet
- Teaching Students With Autism Spectrum DisordersDocument155 pagesTeaching Students With Autism Spectrum DisordersAnonymous Pj6Odj0% (1)
- (Donald E. Super, Charles Super,) OpportunitiesDocument162 pages(Donald E. Super, Charles Super,) OpportunitiesRuthnaomi Vitaloka LiquisaNo ratings yet
- Reflection Papers On Sense and PerceptionDocument2 pagesReflection Papers On Sense and PerceptionAgustin Barretto50% (4)
- Cognitive Load Theory (John Sweller) : Information ProcessingDocument2 pagesCognitive Load Theory (John Sweller) : Information ProcessingNik ZazlealizaNo ratings yet
- Chapter 13 - Goal SettingDocument43 pagesChapter 13 - Goal SettingHANAH LORAINE LAUREANo ratings yet
- Hardiman 2011Document9 pagesHardiman 2011Cristina GutiérrezNo ratings yet
- Characteristics For Observers: Halo Effect Observer DriftDocument1 pageCharacteristics For Observers: Halo Effect Observer DriftHanif PramadanNo ratings yet
- Unit 1: Introduction To Understanding The SelfDocument10 pagesUnit 1: Introduction To Understanding The SelfRosana Celeste CañonNo ratings yet
- Key Concepts of Child and AdolescentDocument59 pagesKey Concepts of Child and Adolescentkm lagangNo ratings yet
- Perceptual ProcessDocument4 pagesPerceptual ProcessDeep K Desai100% (1)
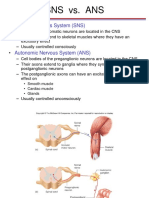
- Sns vs. Ans: Somatic Nervous System (SNS)Document20 pagesSns vs. Ans: Somatic Nervous System (SNS)dave_1128No ratings yet