Download as pdf or txt
You might also like
- Sleep ChartDocument3 pagesSleep ChartCheetah GemmaNo ratings yet
- GBS Case StudyDocument5 pagesGBS Case StudyDelphy Varghese0% (1)
- J Jns 2012 03 027Document9 pagesJ Jns 2012 03 027Juan Jose Eraso OsorioNo ratings yet
- Demencia Vascular 2Document45 pagesDemencia Vascular 21821ainaNo ratings yet
- Cerebral Small Vessel DiseaseDocument11 pagesCerebral Small Vessel Diseaseyousra yatrebNo ratings yet
- Demencia 2 PDFDocument10 pagesDemencia 2 PDFSebastian OrdoñezNo ratings yet
- NIH Public Access: Vascular DementiaDocument23 pagesNIH Public Access: Vascular DementianadaNo ratings yet
- Dementia A Neurodegenerative DisorderDocument16 pagesDementia A Neurodegenerative DisorderRishabh SinghNo ratings yet
- Diabetes, Stroke, and Neuroresilience Looking BeyondDocument21 pagesDiabetes, Stroke, and Neuroresilience Looking BeyondMaudia IndrianiiNo ratings yet
- Review Article: George P. Paraskevas, Vasilios C. Constantinides, Panagiotis G. Paraskevas, Elisabeth KapakiDocument6 pagesReview Article: George P. Paraskevas, Vasilios C. Constantinides, Panagiotis G. Paraskevas, Elisabeth KapakiasclepiuspdfsNo ratings yet
- Fnagi 05 00017Document19 pagesFnagi 05 00017mariamojegNo ratings yet
- Immunity & Ageing: Pathophysiology of Vascular DementiaDocument9 pagesImmunity & Ageing: Pathophysiology of Vascular DementiaMuhammad Cholis HidayatNo ratings yet
- Cerebral Small Vessel Disease: Review ArticleDocument21 pagesCerebral Small Vessel Disease: Review ArticleSiklus AnakNo ratings yet
- Beyond Mild Cognitive Impairment: Vascular Cognitive Impairment, No Dementia (VCIND)Document9 pagesBeyond Mild Cognitive Impairment: Vascular Cognitive Impairment, No Dementia (VCIND)Naresh SharmaNo ratings yet
- Hypertension and DementiaDocument9 pagesHypertension and DementiaJenny LeeNo ratings yet
- Fibrinogen, A Possible Key Player in Alzheimer Õs Disease: M.Cortes-Canteliands - StricklandDocument5 pagesFibrinogen, A Possible Key Player in Alzheimer Õs Disease: M.Cortes-Canteliands - Stricklandwm9105No ratings yet
- DementiaDocument12 pagesDementiaPriscila SelingardiNo ratings yet
- Vascular DementiaDocument8 pagesVascular Dementiaddelindaaa100% (1)
- Vascular Instability and Neurological Morbidity in Sickle Cell Disease: An Integrative Framework.Document21 pagesVascular Instability and Neurological Morbidity in Sickle Cell Disease: An Integrative Framework.Arturo Martí-CarvajalNo ratings yet
- Review: The Pathobiology of Vascular DementiaDocument23 pagesReview: The Pathobiology of Vascular Dementiahiazz17No ratings yet
- The Neuropathology and Cerebrovascular Mechanisms of DementiaDocument15 pagesThe Neuropathology and Cerebrovascular Mechanisms of DementiaRickyRicardoSandyPutraNo ratings yet
- Nasrabady 2018Document10 pagesNasrabady 2018Alix AliNo ratings yet
- Blood Brain Barrier Failure As A Core Mechanism in Cerebral - 2017 - AlzheimerDocument10 pagesBlood Brain Barrier Failure As A Core Mechanism in Cerebral - 2017 - AlzheimerLuis Gerardo Escandon AlcazarNo ratings yet
- Nihms 1698838Document34 pagesNihms 1698838meliaaammeNo ratings yet
- Cognition, Mood and Behavior in CADASILDocument6 pagesCognition, Mood and Behavior in CADASILAna Carolina RibasNo ratings yet
- Deterioro Cognitivo Vascular y Demencia 2016Document9 pagesDeterioro Cognitivo Vascular y Demencia 2016Lalito Manuel Cabellos AcuñaNo ratings yet
- Neuropsychology Dementia ProfilesDocument12 pagesNeuropsychology Dementia ProfilesN SNo ratings yet
- AD Review 2Document14 pagesAD Review 2Emmanuel JacquezNo ratings yet
- Artigo AlzheimerDocument21 pagesArtigo Alzheimerbeatriz assunesNo ratings yet
- The Comorbidity of Alzheimers Disease and Bipolar Disorder and The Potential of Lithium As Drug TherapyDocument16 pagesThe Comorbidity of Alzheimers Disease and Bipolar Disorder and The Potential of Lithium As Drug TherapyAthenaeum Scientific PublishersNo ratings yet
- Stroke and Dementia: Dr. Dr. Suherman, SP.S (K)Document36 pagesStroke and Dementia: Dr. Dr. Suherman, SP.S (K)deri rivanoNo ratings yet
- Neuropatologia AlzheimerDocument27 pagesNeuropatologia AlzheimerSarah JatobáNo ratings yet
- Hypertension-Induced Cognitive Impairment: From Pathophysiology To Public HealthDocument17 pagesHypertension-Induced Cognitive Impairment: From Pathophysiology To Public HealthAulia ayu ningtyasNo ratings yet
- Alzheimer's Disease: Genes, Pathogenesis and Risk PredictionDocument7 pagesAlzheimer's Disease: Genes, Pathogenesis and Risk PredictionWindaNo ratings yet
- Demensia Vaskuler 2Document20 pagesDemensia Vaskuler 2Adam MochtarNo ratings yet
- SDVSDFVDocument12 pagesSDVSDFVmr dexterNo ratings yet
- NIH Public Access: Author ManuscriptDocument9 pagesNIH Public Access: Author ManuscriptNicolás QuintanillaNo ratings yet
- Vascular Dementia: Prevention and Treatment: Catherine Mcveigh Peter PassmoreDocument7 pagesVascular Dementia: Prevention and Treatment: Catherine Mcveigh Peter PassmoreShelia LewisNo ratings yet
- Neuroregeneration FIX BEUTTDocument61 pagesNeuroregeneration FIX BEUTTNurcahyo Tri UtomoNo ratings yet
- Epidemiology, Pathology, and Pathogenesis of Alzheimer Disease - UpToDateDocument31 pagesEpidemiology, Pathology, and Pathogenesis of Alzheimer Disease - UpToDateVolt HakimNo ratings yet
- JURNALDocument10 pagesJURNALLuses Shantia HaryantoNo ratings yet
- Neurological Complications of Sickle Cell DiseaseDocument8 pagesNeurological Complications of Sickle Cell Disease1.thekey.1No ratings yet
- Vascular Cognitive Impairment and Dementia JACC Scientific Expert PanelDocument19 pagesVascular Cognitive Impairment and Dementia JACC Scientific Expert PanelFlavia Ozuna SalinasNo ratings yet
- Clinical Manifestations, Prevention and Treatment of Stroke in WomenDocument6 pagesClinical Manifestations, Prevention and Treatment of Stroke in WomenCentral Asian StudiesNo ratings yet
- Natural Products For Neurodegenerative disorders-SM CommentsDocument34 pagesNatural Products For Neurodegenerative disorders-SM CommentsDipongkar rayNo ratings yet
- Age-Related Immune Alterations and Cerebrovascular in AmmationDocument16 pagesAge-Related Immune Alterations and Cerebrovascular in Ammationzuhdi azzyumar dhiniNo ratings yet
- Cadasil Syndrome: A Case Report With A Literature ReviewDocument4 pagesCadasil Syndrome: A Case Report With A Literature ReviewAna Carolina RibasNo ratings yet
- Circresaha 116 308426 PDFDocument19 pagesCircresaha 116 308426 PDFRahma BachriNo ratings yet
- DNND 37353 Post Stroke Cognitive Impairment and Dementia Prevalence 020514Document7 pagesDNND 37353 Post Stroke Cognitive Impairment and Dementia Prevalence 020514Irham TahkikNo ratings yet
- Reviews: Ageing As A Risk Factor For Neurodegenerative DiseaseDocument17 pagesReviews: Ageing As A Risk Factor For Neurodegenerative DiseaseEla MishraNo ratings yet
- Nihms 1502302Document39 pagesNihms 1502302Мари АлитаNo ratings yet
- Neurological Manifestations of Vascular DementiaDocument5 pagesNeurological Manifestations of Vascular DementiaDumitruAuraNo ratings yet
- Alzheimer Dementia JAMADocument4 pagesAlzheimer Dementia JAMAAna RendónNo ratings yet
- Inflammatory DisordersDocument4 pagesInflammatory DisordersArouetNo ratings yet
- Circresaha 118 313400Document3 pagesCircresaha 118 313400CARDIO 2019No ratings yet
- 8 FullDocument3 pages8 FulladriricaldeNo ratings yet
- Stroke GeneticsDocument13 pagesStroke GeneticsRenato RamonNo ratings yet
- Myocardial Fibrosis in Chagas Disease and Molecules Related To FibrosisDocument7 pagesMyocardial Fibrosis in Chagas Disease and Molecules Related To FibrosisLaura montillaNo ratings yet
- Dichgans M. (2007)Document13 pagesDichgans M. (2007)Rusda AnandaNo ratings yet
- 829 FullDocument11 pages829 FullAsep RiswandiNo ratings yet
- Amyl Oidos IsDocument13 pagesAmyl Oidos IsJose Joey Jumaoas Jr.No ratings yet
- Failure of Deactivation in The Default Mode Network A Trait Marker For SchizophreniaDocument11 pagesFailure of Deactivation in The Default Mode Network A Trait Marker For SchizophreniaJuan Jose Eraso OsorioNo ratings yet
- Connectome and Schizophrenia.Document7 pagesConnectome and Schizophrenia.Juan Jose Eraso OsorioNo ratings yet
- Facial Emotion Processing in Schizophrenia A Non-Specific Neuropsychological DeficitDocument9 pagesFacial Emotion Processing in Schizophrenia A Non-Specific Neuropsychological DeficitJuan Jose Eraso OsorioNo ratings yet
- Association of Formal Thought Disorder in Schizophrenia With Structural Brain Abnormalities in Language-Related Cortical Regions.Document6 pagesAssociation of Formal Thought Disorder in Schizophrenia With Structural Brain Abnormalities in Language-Related Cortical Regions.Juan Jose Eraso OsorioNo ratings yet
- Aproximacion Biopsicosocial A Ttnos SomatomorfosDocument24 pagesAproximacion Biopsicosocial A Ttnos SomatomorfosJuan Jose Eraso OsorioNo ratings yet
- Cuadernillo P-IpgDocument6 pagesCuadernillo P-IpgJuan Jose Eraso OsorioNo ratings yet
- Novel Insight Into Intellectual Disability A RevieDocument9 pagesNovel Insight Into Intellectual Disability A RevieJuan Jose Eraso OsorioNo ratings yet
- Right Care 3. Drivers of Poor Medical CareDocument13 pagesRight Care 3. Drivers of Poor Medical CareJuan Jose Eraso OsorioNo ratings yet
- J Jns 2012 03 027Document9 pagesJ Jns 2012 03 027Juan Jose Eraso OsorioNo ratings yet
- Stroke Compendium: Vascular Cognitive ImpairmentDocument20 pagesStroke Compendium: Vascular Cognitive ImpairmentJuan Jose Eraso OsorioNo ratings yet
- Endoscopic Retrograde Cholangiopancreatography Is.31Document6 pagesEndoscopic Retrograde Cholangiopancreatography Is.31Gefaritza RabbaniNo ratings yet
- NANDA Nursing Diagnosis List 2018Document4 pagesNANDA Nursing Diagnosis List 2018KYLE MITZIE SENGCONo ratings yet
- ABG, CBC, BONE Marrow BiopsyDocument34 pagesABG, CBC, BONE Marrow Biopsymarthonsilva123No ratings yet
- Phototherapy Guidelines-MuneezaDocument8 pagesPhototherapy Guidelines-MuneezaPeki BirdNo ratings yet
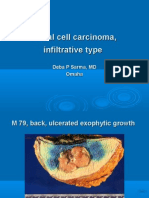
- Basal Cell Carcinoma, Infiltrative TypeDocument8 pagesBasal Cell Carcinoma, Infiltrative TypeDeba P SarmaNo ratings yet
- Nursing Research Position StatementDocument3 pagesNursing Research Position StatementFranck Duprey MvogoNo ratings yet
- In Vitro Prediction of in Vivo Bioavailability andDocument8 pagesIn Vitro Prediction of in Vivo Bioavailability andMayang OliviaNo ratings yet
- Optimized Ear Acupuncture (EAF)Document9 pagesOptimized Ear Acupuncture (EAF)PhilNo ratings yet
- Otitis Media and ExternaDocument22 pagesOtitis Media and ExternaLhanz CayingNo ratings yet
- Case Study On HEAD INJURYDocument11 pagesCase Study On HEAD INJURYanimesh pandaNo ratings yet
- TWSTRS Severity ScaleDocument2 pagesTWSTRS Severity ScaleErri PratamaNo ratings yet
- Wolfis Law Letter PrintDocument1 pageWolfis Law Letter PrintSatyam PatelNo ratings yet
- Pathology Week-14Document9 pagesPathology Week-14taysi tafriNo ratings yet
- Epidural Anesthesia: Grace G. Enriquez, MD, DPBADocument38 pagesEpidural Anesthesia: Grace G. Enriquez, MD, DPBANikki Tuazon BartolomeNo ratings yet
- 93 42 Eec - enDocument2 pages93 42 Eec - enthrewaway100% (1)
- Case Study Med SurgDocument2 pagesCase Study Med SurgKayla NapolitanoNo ratings yet
- Heat Illness Prevention Training 20170328Document19 pagesHeat Illness Prevention Training 20170328CL1P0No ratings yet
- Flight DocumentsDocument1 pageFlight DocumentsNguyen Thang LoiNo ratings yet
- Drug Study AmoxicillinDocument3 pagesDrug Study AmoxicillinJanzelvine Lee MontenegroNo ratings yet
- Pharmacy Program BluecrossDocument46 pagesPharmacy Program BluecrossSarah WeeksNo ratings yet
- SeminarDocument34 pagesSeminarMuhammad Faiz Satria WibowoNo ratings yet
- Tree of LifeDocument909 pagesTree of LifeMariza D. KabamaruNo ratings yet
- Grid For Prof Devel PlanDocument1 pageGrid For Prof Devel Planapi-302124528No ratings yet
- Lindsley Et Al-2017-The Cochrane Library - Sup-1Document3 pagesLindsley Et Al-2017-The Cochrane Library - Sup-1Andreina VeraNo ratings yet
- DKA Handout1Document59 pagesDKA Handout1aleNo ratings yet
- Hospital ManagementDocument3 pagesHospital ManagementFebriciaKristinaNo ratings yet
- Intestinal Ischemia in Neonates and Children: ReviewDocument5 pagesIntestinal Ischemia in Neonates and Children: Reviewriri siahaanNo ratings yet
- OxytocicsDocument36 pagesOxytocicsNovitaNo ratings yet