
Pharmaceutical Techonolgy
Pharmaceutical Techonolgy
You might also like
- Astm E84Document19 pagesAstm E84Juan Felipe Grisales González100% (1)
- Clmd4a Scieg4.PubDocument40 pagesClmd4a Scieg4.PubAnne YouNo ratings yet
- Caprine Milk Oligos NZ Saanen Goat Cheese Whey Composition Enrichment Thum 2015Document8 pagesCaprine Milk Oligos NZ Saanen Goat Cheese Whey Composition Enrichment Thum 2015Don OtterNo ratings yet
- Hadrová 2012Document11 pagesHadrová 2012leandro mendesNo ratings yet
- Nutrients 10 00262Document15 pagesNutrients 10 00262Sally Salina FarahNo ratings yet
- Modifications Induced by Controlled Storage Conditions On Whey Protein ConcentratesDocument32 pagesModifications Induced by Controlled Storage Conditions On Whey Protein ConcentratesRed riotNo ratings yet
- Journal of Food Biochemistry - 2022 - Iram - in Silico Identification of Antidiabetic and Hypotensive Potential BioactiveDocument16 pagesJournal of Food Biochemistry - 2022 - Iram - in Silico Identification of Antidiabetic and Hypotensive Potential BioactiveAnia KononiukNo ratings yet
- Biological Evaluation of Fermented Camel Milk Drinkable Like Soup Fortified With Different CerealsDocument7 pagesBiological Evaluation of Fermented Camel Milk Drinkable Like Soup Fortified With Different CerealsMarwa ElgendyNo ratings yet
- Atrovas Dan Feno FibrateDocument9 pagesAtrovas Dan Feno FibrateGalih Antona SinatriaNo ratings yet
- Whey Protein Concentrate Hydrolyzed by Microbial Protease: Process Optimization and Evaluation of Its Dipeptidyl Peptidase Inhibitory ActivityDocument13 pagesWhey Protein Concentrate Hydrolyzed by Microbial Protease: Process Optimization and Evaluation of Its Dipeptidyl Peptidase Inhibitory ActivityprasannaNo ratings yet
- Literature Review On Microbial Evaluation of Cow MilkDocument5 pagesLiterature Review On Microbial Evaluation of Cow MilkygivrcxgfNo ratings yet
- Effect of Linseed Oil2 PDFDocument12 pagesEffect of Linseed Oil2 PDFSergio TunNo ratings yet
- Tsiplakou Et Al., 2017a-LDocument30 pagesTsiplakou Et Al., 2017a-LLuis Alberto Canul KuNo ratings yet
- Yu2019 - Quanification of Nervonic Acid in Human MilkDocument14 pagesYu2019 - Quanification of Nervonic Acid in Human MilkOlgaNo ratings yet
- Antihypertensive Effect of Bioactive Peptides Produced by Protease-Facilitated Lactic Acid Fermentation of MilkDocument7 pagesAntihypertensive Effect of Bioactive Peptides Produced by Protease-Facilitated Lactic Acid Fermentation of MilkmanhalNo ratings yet
- Accepted Version: Theriogenology. Changes Resulting From The Publishing Process, Such As Peer ReviewDocument30 pagesAccepted Version: Theriogenology. Changes Resulting From The Publishing Process, Such As Peer Reviewanahh ramakNo ratings yet
- Effect of Ferulic Acid On Cholesterol Synthesis Serum Lipids and Hepatotoxicity in Atherogenic RatsDocument7 pagesEffect of Ferulic Acid On Cholesterol Synthesis Serum Lipids and Hepatotoxicity in Atherogenic RatsvinayNo ratings yet
- jfnr201711 PDFDocument8 pagesjfnr201711 PDFAyu Puspita FebrindariNo ratings yet
- 1 s2.0 S0022030212002044 MainDocument19 pages1 s2.0 S0022030212002044 MainFriska YulandaNo ratings yet
- Analysis of Milk Fats From Different MammalianDocument11 pagesAnalysis of Milk Fats From Different MammalianPavel RoutNo ratings yet
- Research Article: Egg White Hydrolysate Retains The Nutritional Value of Proteins and Is Quickly Absorbed in RatsDocument7 pagesResearch Article: Egg White Hydrolysate Retains The Nutritional Value of Proteins and Is Quickly Absorbed in Ratsnah indoNo ratings yet
- 1 Ijapbcraug20171Document6 pages1 Ijapbcraug20171TJPRC PublicationsNo ratings yet
- Effect of Bitter Gourd (Momordica Charantia) On Glycaemic Status in Streptozotocin Induced Diabetic RatsDocument5 pagesEffect of Bitter Gourd (Momordica Charantia) On Glycaemic Status in Streptozotocin Induced Diabetic Ratsjeje pontillasNo ratings yet
- Yield and Composition of Milk From Dairy Cows Fed Diets Containing Cashew NutsDocument8 pagesYield and Composition of Milk From Dairy Cows Fed Diets Containing Cashew Nutsonurb90No ratings yet
- 451-Article Text-487-1-10-20151017Document9 pages451-Article Text-487-1-10-20151017Maulika TazkiaNo ratings yet
- 751149v1 Full PDFDocument30 pages751149v1 Full PDFMaria AspriNo ratings yet
- Accepted Manuscript: Animal Feed Science and TechnologyDocument53 pagesAccepted Manuscript: Animal Feed Science and TechnologyGabriela Casarotto DanielNo ratings yet
- High Pressure Homogenization Hemp MilkDocument8 pagesHigh Pressure Homogenization Hemp Milkmarcela cadenaNo ratings yet
- J. Lipid Res. 1982 Denning 584 96Document13 pagesJ. Lipid Res. 1982 Denning 584 96stbudaNo ratings yet
- Biomedicine & Pharmacotherapy: SciencedirectDocument11 pagesBiomedicine & Pharmacotherapy: SciencedirectAna Raíza OliveiraNo ratings yet
- 1 s2.0 S0742841396002290 MainDocument8 pages1 s2.0 S0742841396002290 MainEuclides DamiãoNo ratings yet
- Soybean Whey Enhance Mineral Balance and Caecal Fermentation in RatsDocument9 pagesSoybean Whey Enhance Mineral Balance and Caecal Fermentation in RatsThalia KarampasiNo ratings yet
- ArticleDocument7 pagesArticleAline Lopes NascimetoNo ratings yet
- Peptides: John P. Gleeson, Joanne Heade, Sinéad M. Ryan, David J. BraydenDocument7 pagesPeptides: John P. Gleeson, Joanne Heade, Sinéad M. Ryan, David J. Braydensuci apsari pebriantiNo ratings yet
- In Vitro Digestion of Red Deer (Cervus Elaphus) and Cow (Bos Taurus) MilkDocument8 pagesIn Vitro Digestion of Red Deer (Cervus Elaphus) and Cow (Bos Taurus) MilkEmmanuel Mpaliye MpaliyeNo ratings yet
- J ANIM SCI-1984-Wooden-1529-35Document8 pagesJ ANIM SCI-1984-Wooden-1529-35sidakanNo ratings yet
- Hydrolsis NyamplungDocument7 pagesHydrolsis NyamplungVenny AgustinNo ratings yet
- 1 s2.0 S0963996923003812 MainDocument9 pages1 s2.0 S0963996923003812 Mainjair barruetaNo ratings yet
- Effect of Solid-State Fermentation On Proximate Composition, Anti-Nutritional Factor, Microbiological and Functional Properties of Lupin FlourDocument28 pagesEffect of Solid-State Fermentation On Proximate Composition, Anti-Nutritional Factor, Microbiological and Functional Properties of Lupin FlourAkbar AJNo ratings yet
- Effects of Exogenous Lactate On Lipid, Protein, and Glucose Metabolism (Pedersen Et Al., 2024)Document40 pagesEffects of Exogenous Lactate On Lipid, Protein, and Glucose Metabolism (Pedersen Et Al., 2024)Marcelo TrNo ratings yet
- 2017 Feeding of Palm Oil Fatty Acids or Rapeseed Oil ThroughoutDocument14 pages2017 Feeding of Palm Oil Fatty Acids or Rapeseed Oil ThroughoutJeferson Lopez RomeroNo ratings yet
- Changes of Isoflavones During Processing of Soy Protein IsolatesDocument5 pagesChanges of Isoflavones During Processing of Soy Protein IsolatesDinh DungNo ratings yet
- Effect of The Dietary By-Product Grape Seed Cake On Performances and Carcass Quality of TOPIGS PigsDocument10 pagesEffect of The Dietary By-Product Grape Seed Cake On Performances and Carcass Quality of TOPIGS PigsmariaNo ratings yet
- Acidophilus and Lactobacillus Casei of Human Intestinal OriginDocument6 pagesAcidophilus and Lactobacillus Casei of Human Intestinal Originmmoradi55No ratings yet
- 2015 Article 263 (Articulo2)Document31 pages2015 Article 263 (Articulo2)GRACIELANo ratings yet
- 1 s2.0 S0023643821017345 MainDocument8 pages1 s2.0 S0023643821017345 MainRochman SyahNo ratings yet
- J. AOAC Int 2002, Vol 85 (5), Pages 1090-1095Document6 pagesJ. AOAC Int 2002, Vol 85 (5), Pages 1090-1095Thanh Tâm TrầnNo ratings yet
- Nutritional Importance of OrangesDocument18 pagesNutritional Importance of OrangesVarsha RaniNo ratings yet
- Effect of Different Factors On Goat MilkDocument6 pagesEffect of Different Factors On Goat MilkMohammed Mizhab K MNo ratings yet
- CLA LecheDocument8 pagesCLA LecheWalter Vega FernándezNo ratings yet
- Omega-3/6 Fatty Acids: Alternative Sources of Production: Owen P. Ward, Ajay SinghDocument26 pagesOmega-3/6 Fatty Acids: Alternative Sources of Production: Owen P. Ward, Ajay SinghVictor Muñoz GodoyNo ratings yet
- Enzymatic Hydrolysis of Wheat Gluten by Proteases and Properties of The Resulting HydrolysatesDocument5 pagesEnzymatic Hydrolysis of Wheat Gluten by Proteases and Properties of The Resulting HydrolysatesPadmini RajuNo ratings yet
- Effect of Wakame On Non Alcoholic Fatty LiverDocument14 pagesEffect of Wakame On Non Alcoholic Fatty LiverCătălina RădulescuNo ratings yet
- Rice Bran Reduces Weight Gain and Modulates Lipid Metabolism in Rats With High-Energy-Diet-Induced ObesityDocument10 pagesRice Bran Reduces Weight Gain and Modulates Lipid Metabolism in Rats With High-Energy-Diet-Induced ObesityXavier Alvaro Quille MamaniNo ratings yet
- JCTM 2024 59 1 2 23-22 pp15-22Document8 pagesJCTM 2024 59 1 2 23-22 pp15-22UNSNo ratings yet
- Spent Coffee GroundsDocument9 pagesSpent Coffee GroundsNishadh NishNo ratings yet
- Influence of Flaxseed Components On Fermented DairyDocument6 pagesInfluence of Flaxseed Components On Fermented DairyEl-Khansa DEBBACHENo ratings yet
- John K. Mathai, Yanhong Liu and Hans H. SteinDocument10 pagesJohn K. Mathai, Yanhong Liu and Hans H. Steinaryadave20No ratings yet
- 1472 6882 2 1 PDFDocument7 pages1472 6882 2 1 PDFThiago NunesNo ratings yet
- Effect of Pleurotus Tuber-Regium Polysaccharides Supplementation On The Progression of Diabetes Complications in Obese-Diabetic RatsDocument11 pagesEffect of Pleurotus Tuber-Regium Polysaccharides Supplementation On The Progression of Diabetes Complications in Obese-Diabetic RatsAlmaNo ratings yet
- Cytoskeleton 1 PDFDocument4 pagesCytoskeleton 1 PDFPortgas AceNo ratings yet
- MSDS - 901 S3 10WDocument5 pagesMSDS - 901 S3 10WGirish SanghiNo ratings yet
- Bioplastic From Banana Peel Research PaperDocument8 pagesBioplastic From Banana Peel Research Papergvytgh3b100% (2)
- Tot Lunaria KT 46 MsdsDocument4 pagesTot Lunaria KT 46 Msdsnoar anwarNo ratings yet
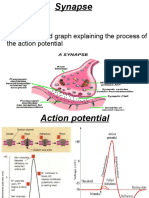
- Synapse 1214834359710231 8 PDFDocument11 pagesSynapse 1214834359710231 8 PDFMuhammad AbdullahNo ratings yet
- Clariant Flyer ReforMax LDP Plus 201702 enDocument2 pagesClariant Flyer ReforMax LDP Plus 201702 enNguyễn Văn MinhNo ratings yet
- Summative Examination Cookery SY 2020-2021 I. Multiple Choice. Select The Best AnswerDocument2 pagesSummative Examination Cookery SY 2020-2021 I. Multiple Choice. Select The Best AnswerZyreane FernandezNo ratings yet
- RYYQ-T, RYMQ-T, RXYQ-T, RYYQ-T8 - 4PEN370475-1C - 2016 - 11 - Installer and User Reference Guide - EnglishDocument92 pagesRYYQ-T, RYMQ-T, RXYQ-T, RYYQ-T8 - 4PEN370475-1C - 2016 - 11 - Installer and User Reference Guide - EnglishenduNo ratings yet
- Doe4 PDFDocument11 pagesDoe4 PDFlamthanh87No ratings yet
- Preventol® CMK DisinfectionDocument4 pagesPreventol® CMK DisinfectionEdinson Javier Cuello CastroNo ratings yet
- Production of Ceramic Foam Filters For Molten MetaDocument5 pagesProduction of Ceramic Foam Filters For Molten MetaVBCRC LABNo ratings yet
- Investigation of The Potential of Cyperus AlternifoliusDocument8 pagesInvestigation of The Potential of Cyperus AlternifoliusCt Kamariah Md SaatNo ratings yet
- Report RobotDocument10 pagesReport RobotAimy SufeaNo ratings yet
- Linezolid Finished Testing MethodDocument4 pagesLinezolid Finished Testing MethodLife PearlNo ratings yet
- Is 14428 1997 PDFDocument11 pagesIs 14428 1997 PDFdurg0706No ratings yet
- 159 B EutecRodDocument4 pages159 B EutecRodMohamed AdelNo ratings yet
- Test 1 - Redox ReactionsDocument5 pagesTest 1 - Redox Reactionsaliesha cassasolaNo ratings yet
- Plant Biotechnology Lecture 2 10-10-22Document23 pagesPlant Biotechnology Lecture 2 10-10-22Youssef AbdelhayNo ratings yet
- Environmental Protection Agency (EPA) (Timeline and Role in Society)Document25 pagesEnvironmental Protection Agency (EPA) (Timeline and Role in Society)Abeera MalikNo ratings yet
- DiagenesisDocument2 pagesDiagenesisMadhurjya PhukanNo ratings yet
- Msds Bezaprint Red CC-GDocument9 pagesMsds Bezaprint Red CC-GunisourcceeNo ratings yet
- Teromatec 400 English Manual PDFDocument24 pagesTeromatec 400 English Manual PDFex-2156100% (1)
- Ujian Akhir Semester Elusiasi Struktur (Senyawa Organik) T.A: 2019-2020 GenapDocument11 pagesUjian Akhir Semester Elusiasi Struktur (Senyawa Organik) T.A: 2019-2020 GenapArianto UmarNo ratings yet
- Chemistry (5070) : Summer HomeworkDocument19 pagesChemistry (5070) : Summer HomeworkAmeer HamzaNo ratings yet
- ESA Sample Specification Herbs and Spices (2003)Document3 pagesESA Sample Specification Herbs and Spices (2003)Mumtaz QcNo ratings yet
- Legit Multivector Review and Training Center Online Self-Assessment ExaminationDocument6 pagesLegit Multivector Review and Training Center Online Self-Assessment ExaminationJunnar Jay Abaño100% (1)
- WFP-0000020229 - Argentina MaizeDocument3 pagesWFP-0000020229 - Argentina MaizeThuat NguyenNo ratings yet
- Sheng 2014Document19 pagesSheng 201456962645No ratings yet
- CH 32Document16 pagesCH 32dkhatri01No ratings yet
























































































Download as pdf or txt
You might also like
- Astm E84Document19 pagesAstm E84Juan Felipe Grisales González100% (1)
- Clmd4a Scieg4.PubDocument40 pagesClmd4a Scieg4.PubAnne YouNo ratings yet
- Caprine Milk Oligos NZ Saanen Goat Cheese Whey Composition Enrichment Thum 2015Document8 pagesCaprine Milk Oligos NZ Saanen Goat Cheese Whey Composition Enrichment Thum 2015Don OtterNo ratings yet
- Hadrová 2012Document11 pagesHadrová 2012leandro mendesNo ratings yet
- Nutrients 10 00262Document15 pagesNutrients 10 00262Sally Salina FarahNo ratings yet
- Modifications Induced by Controlled Storage Conditions On Whey Protein ConcentratesDocument32 pagesModifications Induced by Controlled Storage Conditions On Whey Protein ConcentratesRed riotNo ratings yet
- Journal of Food Biochemistry - 2022 - Iram - in Silico Identification of Antidiabetic and Hypotensive Potential BioactiveDocument16 pagesJournal of Food Biochemistry - 2022 - Iram - in Silico Identification of Antidiabetic and Hypotensive Potential BioactiveAnia KononiukNo ratings yet
- Biological Evaluation of Fermented Camel Milk Drinkable Like Soup Fortified With Different CerealsDocument7 pagesBiological Evaluation of Fermented Camel Milk Drinkable Like Soup Fortified With Different CerealsMarwa ElgendyNo ratings yet
- Atrovas Dan Feno FibrateDocument9 pagesAtrovas Dan Feno FibrateGalih Antona SinatriaNo ratings yet
- Whey Protein Concentrate Hydrolyzed by Microbial Protease: Process Optimization and Evaluation of Its Dipeptidyl Peptidase Inhibitory ActivityDocument13 pagesWhey Protein Concentrate Hydrolyzed by Microbial Protease: Process Optimization and Evaluation of Its Dipeptidyl Peptidase Inhibitory ActivityprasannaNo ratings yet
- Literature Review On Microbial Evaluation of Cow MilkDocument5 pagesLiterature Review On Microbial Evaluation of Cow MilkygivrcxgfNo ratings yet
- Effect of Linseed Oil2 PDFDocument12 pagesEffect of Linseed Oil2 PDFSergio TunNo ratings yet
- Tsiplakou Et Al., 2017a-LDocument30 pagesTsiplakou Et Al., 2017a-LLuis Alberto Canul KuNo ratings yet
- Yu2019 - Quanification of Nervonic Acid in Human MilkDocument14 pagesYu2019 - Quanification of Nervonic Acid in Human MilkOlgaNo ratings yet
- Antihypertensive Effect of Bioactive Peptides Produced by Protease-Facilitated Lactic Acid Fermentation of MilkDocument7 pagesAntihypertensive Effect of Bioactive Peptides Produced by Protease-Facilitated Lactic Acid Fermentation of MilkmanhalNo ratings yet
- Accepted Version: Theriogenology. Changes Resulting From The Publishing Process, Such As Peer ReviewDocument30 pagesAccepted Version: Theriogenology. Changes Resulting From The Publishing Process, Such As Peer Reviewanahh ramakNo ratings yet
- Effect of Ferulic Acid On Cholesterol Synthesis Serum Lipids and Hepatotoxicity in Atherogenic RatsDocument7 pagesEffect of Ferulic Acid On Cholesterol Synthesis Serum Lipids and Hepatotoxicity in Atherogenic RatsvinayNo ratings yet
- jfnr201711 PDFDocument8 pagesjfnr201711 PDFAyu Puspita FebrindariNo ratings yet
- 1 s2.0 S0022030212002044 MainDocument19 pages1 s2.0 S0022030212002044 MainFriska YulandaNo ratings yet
- Analysis of Milk Fats From Different MammalianDocument11 pagesAnalysis of Milk Fats From Different MammalianPavel RoutNo ratings yet
- Research Article: Egg White Hydrolysate Retains The Nutritional Value of Proteins and Is Quickly Absorbed in RatsDocument7 pagesResearch Article: Egg White Hydrolysate Retains The Nutritional Value of Proteins and Is Quickly Absorbed in Ratsnah indoNo ratings yet
- 1 Ijapbcraug20171Document6 pages1 Ijapbcraug20171TJPRC PublicationsNo ratings yet
- Effect of Bitter Gourd (Momordica Charantia) On Glycaemic Status in Streptozotocin Induced Diabetic RatsDocument5 pagesEffect of Bitter Gourd (Momordica Charantia) On Glycaemic Status in Streptozotocin Induced Diabetic Ratsjeje pontillasNo ratings yet
- Yield and Composition of Milk From Dairy Cows Fed Diets Containing Cashew NutsDocument8 pagesYield and Composition of Milk From Dairy Cows Fed Diets Containing Cashew Nutsonurb90No ratings yet
- 451-Article Text-487-1-10-20151017Document9 pages451-Article Text-487-1-10-20151017Maulika TazkiaNo ratings yet
- 751149v1 Full PDFDocument30 pages751149v1 Full PDFMaria AspriNo ratings yet
- Accepted Manuscript: Animal Feed Science and TechnologyDocument53 pagesAccepted Manuscript: Animal Feed Science and TechnologyGabriela Casarotto DanielNo ratings yet
- High Pressure Homogenization Hemp MilkDocument8 pagesHigh Pressure Homogenization Hemp Milkmarcela cadenaNo ratings yet
- J. Lipid Res. 1982 Denning 584 96Document13 pagesJ. Lipid Res. 1982 Denning 584 96stbudaNo ratings yet
- Biomedicine & Pharmacotherapy: SciencedirectDocument11 pagesBiomedicine & Pharmacotherapy: SciencedirectAna Raíza OliveiraNo ratings yet
- 1 s2.0 S0742841396002290 MainDocument8 pages1 s2.0 S0742841396002290 MainEuclides DamiãoNo ratings yet
- Soybean Whey Enhance Mineral Balance and Caecal Fermentation in RatsDocument9 pagesSoybean Whey Enhance Mineral Balance and Caecal Fermentation in RatsThalia KarampasiNo ratings yet
- ArticleDocument7 pagesArticleAline Lopes NascimetoNo ratings yet
- Peptides: John P. Gleeson, Joanne Heade, Sinéad M. Ryan, David J. BraydenDocument7 pagesPeptides: John P. Gleeson, Joanne Heade, Sinéad M. Ryan, David J. Braydensuci apsari pebriantiNo ratings yet
- In Vitro Digestion of Red Deer (Cervus Elaphus) and Cow (Bos Taurus) MilkDocument8 pagesIn Vitro Digestion of Red Deer (Cervus Elaphus) and Cow (Bos Taurus) MilkEmmanuel Mpaliye MpaliyeNo ratings yet
- J ANIM SCI-1984-Wooden-1529-35Document8 pagesJ ANIM SCI-1984-Wooden-1529-35sidakanNo ratings yet
- Hydrolsis NyamplungDocument7 pagesHydrolsis NyamplungVenny AgustinNo ratings yet
- 1 s2.0 S0963996923003812 MainDocument9 pages1 s2.0 S0963996923003812 Mainjair barruetaNo ratings yet
- Effect of Solid-State Fermentation On Proximate Composition, Anti-Nutritional Factor, Microbiological and Functional Properties of Lupin FlourDocument28 pagesEffect of Solid-State Fermentation On Proximate Composition, Anti-Nutritional Factor, Microbiological and Functional Properties of Lupin FlourAkbar AJNo ratings yet
- Effects of Exogenous Lactate On Lipid, Protein, and Glucose Metabolism (Pedersen Et Al., 2024)Document40 pagesEffects of Exogenous Lactate On Lipid, Protein, and Glucose Metabolism (Pedersen Et Al., 2024)Marcelo TrNo ratings yet
- 2017 Feeding of Palm Oil Fatty Acids or Rapeseed Oil ThroughoutDocument14 pages2017 Feeding of Palm Oil Fatty Acids or Rapeseed Oil ThroughoutJeferson Lopez RomeroNo ratings yet
- Changes of Isoflavones During Processing of Soy Protein IsolatesDocument5 pagesChanges of Isoflavones During Processing of Soy Protein IsolatesDinh DungNo ratings yet
- Effect of The Dietary By-Product Grape Seed Cake On Performances and Carcass Quality of TOPIGS PigsDocument10 pagesEffect of The Dietary By-Product Grape Seed Cake On Performances and Carcass Quality of TOPIGS PigsmariaNo ratings yet
- Acidophilus and Lactobacillus Casei of Human Intestinal OriginDocument6 pagesAcidophilus and Lactobacillus Casei of Human Intestinal Originmmoradi55No ratings yet
- 2015 Article 263 (Articulo2)Document31 pages2015 Article 263 (Articulo2)GRACIELANo ratings yet
- 1 s2.0 S0023643821017345 MainDocument8 pages1 s2.0 S0023643821017345 MainRochman SyahNo ratings yet
- J. AOAC Int 2002, Vol 85 (5), Pages 1090-1095Document6 pagesJ. AOAC Int 2002, Vol 85 (5), Pages 1090-1095Thanh Tâm TrầnNo ratings yet
- Nutritional Importance of OrangesDocument18 pagesNutritional Importance of OrangesVarsha RaniNo ratings yet
- Effect of Different Factors On Goat MilkDocument6 pagesEffect of Different Factors On Goat MilkMohammed Mizhab K MNo ratings yet
- CLA LecheDocument8 pagesCLA LecheWalter Vega FernándezNo ratings yet
- Omega-3/6 Fatty Acids: Alternative Sources of Production: Owen P. Ward, Ajay SinghDocument26 pagesOmega-3/6 Fatty Acids: Alternative Sources of Production: Owen P. Ward, Ajay SinghVictor Muñoz GodoyNo ratings yet
- Enzymatic Hydrolysis of Wheat Gluten by Proteases and Properties of The Resulting HydrolysatesDocument5 pagesEnzymatic Hydrolysis of Wheat Gluten by Proteases and Properties of The Resulting HydrolysatesPadmini RajuNo ratings yet
- Effect of Wakame On Non Alcoholic Fatty LiverDocument14 pagesEffect of Wakame On Non Alcoholic Fatty LiverCătălina RădulescuNo ratings yet
- Rice Bran Reduces Weight Gain and Modulates Lipid Metabolism in Rats With High-Energy-Diet-Induced ObesityDocument10 pagesRice Bran Reduces Weight Gain and Modulates Lipid Metabolism in Rats With High-Energy-Diet-Induced ObesityXavier Alvaro Quille MamaniNo ratings yet
- JCTM 2024 59 1 2 23-22 pp15-22Document8 pagesJCTM 2024 59 1 2 23-22 pp15-22UNSNo ratings yet
- Spent Coffee GroundsDocument9 pagesSpent Coffee GroundsNishadh NishNo ratings yet
- Influence of Flaxseed Components On Fermented DairyDocument6 pagesInfluence of Flaxseed Components On Fermented DairyEl-Khansa DEBBACHENo ratings yet
- John K. Mathai, Yanhong Liu and Hans H. SteinDocument10 pagesJohn K. Mathai, Yanhong Liu and Hans H. Steinaryadave20No ratings yet
- 1472 6882 2 1 PDFDocument7 pages1472 6882 2 1 PDFThiago NunesNo ratings yet
- Effect of Pleurotus Tuber-Regium Polysaccharides Supplementation On The Progression of Diabetes Complications in Obese-Diabetic RatsDocument11 pagesEffect of Pleurotus Tuber-Regium Polysaccharides Supplementation On The Progression of Diabetes Complications in Obese-Diabetic RatsAlmaNo ratings yet
- Cytoskeleton 1 PDFDocument4 pagesCytoskeleton 1 PDFPortgas AceNo ratings yet
- MSDS - 901 S3 10WDocument5 pagesMSDS - 901 S3 10WGirish SanghiNo ratings yet
- Bioplastic From Banana Peel Research PaperDocument8 pagesBioplastic From Banana Peel Research Papergvytgh3b100% (2)
- Tot Lunaria KT 46 MsdsDocument4 pagesTot Lunaria KT 46 Msdsnoar anwarNo ratings yet
- Synapse 1214834359710231 8 PDFDocument11 pagesSynapse 1214834359710231 8 PDFMuhammad AbdullahNo ratings yet
- Clariant Flyer ReforMax LDP Plus 201702 enDocument2 pagesClariant Flyer ReforMax LDP Plus 201702 enNguyễn Văn MinhNo ratings yet
- Summative Examination Cookery SY 2020-2021 I. Multiple Choice. Select The Best AnswerDocument2 pagesSummative Examination Cookery SY 2020-2021 I. Multiple Choice. Select The Best AnswerZyreane FernandezNo ratings yet
- RYYQ-T, RYMQ-T, RXYQ-T, RYYQ-T8 - 4PEN370475-1C - 2016 - 11 - Installer and User Reference Guide - EnglishDocument92 pagesRYYQ-T, RYMQ-T, RXYQ-T, RYYQ-T8 - 4PEN370475-1C - 2016 - 11 - Installer and User Reference Guide - EnglishenduNo ratings yet
- Doe4 PDFDocument11 pagesDoe4 PDFlamthanh87No ratings yet
- Preventol® CMK DisinfectionDocument4 pagesPreventol® CMK DisinfectionEdinson Javier Cuello CastroNo ratings yet
- Production of Ceramic Foam Filters For Molten MetaDocument5 pagesProduction of Ceramic Foam Filters For Molten MetaVBCRC LABNo ratings yet
- Investigation of The Potential of Cyperus AlternifoliusDocument8 pagesInvestigation of The Potential of Cyperus AlternifoliusCt Kamariah Md SaatNo ratings yet
- Report RobotDocument10 pagesReport RobotAimy SufeaNo ratings yet
- Linezolid Finished Testing MethodDocument4 pagesLinezolid Finished Testing MethodLife PearlNo ratings yet
- Is 14428 1997 PDFDocument11 pagesIs 14428 1997 PDFdurg0706No ratings yet
- 159 B EutecRodDocument4 pages159 B EutecRodMohamed AdelNo ratings yet
- Test 1 - Redox ReactionsDocument5 pagesTest 1 - Redox Reactionsaliesha cassasolaNo ratings yet
- Plant Biotechnology Lecture 2 10-10-22Document23 pagesPlant Biotechnology Lecture 2 10-10-22Youssef AbdelhayNo ratings yet
- Environmental Protection Agency (EPA) (Timeline and Role in Society)Document25 pagesEnvironmental Protection Agency (EPA) (Timeline and Role in Society)Abeera MalikNo ratings yet
- DiagenesisDocument2 pagesDiagenesisMadhurjya PhukanNo ratings yet
- Msds Bezaprint Red CC-GDocument9 pagesMsds Bezaprint Red CC-GunisourcceeNo ratings yet
- Teromatec 400 English Manual PDFDocument24 pagesTeromatec 400 English Manual PDFex-2156100% (1)
- Ujian Akhir Semester Elusiasi Struktur (Senyawa Organik) T.A: 2019-2020 GenapDocument11 pagesUjian Akhir Semester Elusiasi Struktur (Senyawa Organik) T.A: 2019-2020 GenapArianto UmarNo ratings yet
- Chemistry (5070) : Summer HomeworkDocument19 pagesChemistry (5070) : Summer HomeworkAmeer HamzaNo ratings yet
- ESA Sample Specification Herbs and Spices (2003)Document3 pagesESA Sample Specification Herbs and Spices (2003)Mumtaz QcNo ratings yet
- Legit Multivector Review and Training Center Online Self-Assessment ExaminationDocument6 pagesLegit Multivector Review and Training Center Online Self-Assessment ExaminationJunnar Jay Abaño100% (1)
- WFP-0000020229 - Argentina MaizeDocument3 pagesWFP-0000020229 - Argentina MaizeThuat NguyenNo ratings yet
- Sheng 2014Document19 pagesSheng 201456962645No ratings yet
- CH 32Document16 pagesCH 32dkhatri01No ratings yet