Download as pdf or txt
You might also like
- 3 Spinal Cord Injury Nursing Care PlanDocument9 pages3 Spinal Cord Injury Nursing Care PlanCyrus De Asis88% (8)
- 2F 5 Radiation Risk AssessmentDocument4 pages2F 5 Radiation Risk AssessmentNurul SyaheerahNo ratings yet
- Biol 235 Assignment-3Document21 pagesBiol 235 Assignment-3Kiron GonidisNo ratings yet
- Operating Room ProcedureDocument4 pagesOperating Room ProcedureROxanne S. RendonNo ratings yet
- Impact of Reduced Intensity Conditioning Allogeneic Stem Cell Transplantation On Women S Fertility 2013 Clinical Lymphoma Myeloma and LeukemiaDocument7 pagesImpact of Reduced Intensity Conditioning Allogeneic Stem Cell Transplantation On Women S Fertility 2013 Clinical Lymphoma Myeloma and LeukemiafujimeisterNo ratings yet
- Geerts 2009Document9 pagesGeerts 2009Berry BancinNo ratings yet
- Molecular Carcinogenesis Endometrium PDFDocument7 pagesMolecular Carcinogenesis Endometrium PDFMelati HasnailNo ratings yet
- Clinicopathological Study of Metallothionein Immunohistochemical Expression, in Benign, Borderline and Malignant Ovarian Epithelial TumorsDocument7 pagesClinicopathological Study of Metallothionein Immunohistochemical Expression, in Benign, Borderline and Malignant Ovarian Epithelial TumorsNajmul HasanNo ratings yet
- CASE REPORT - CA Tiroid Aldi - ProofreadDocument11 pagesCASE REPORT - CA Tiroid Aldi - ProofreadAldy BimaNo ratings yet
- Endometriosis and The Neoplastic Process JurnalDocument12 pagesEndometriosis and The Neoplastic Process JurnalDhe'chi Sudah Tak Tul'lalitNo ratings yet
- Non-Epithelial Ovarian Cancers: Risk ManagementDocument3 pagesNon-Epithelial Ovarian Cancers: Risk ManagementAngga PrabawaNo ratings yet
- BPJ Vol 14 No 4 P 1993-2005Document13 pagesBPJ Vol 14 No 4 P 1993-2005Dr. Shraddha YadavNo ratings yet
- Epithelial To Mesenchymal Transition and Cell Biology of Molecular Regulation in Endometrial CarcinogenesisDocument17 pagesEpithelial To Mesenchymal Transition and Cell Biology of Molecular Regulation in Endometrial CarcinogenesisMelati HasnailNo ratings yet
- CancerDocument26 pagesCancerAnonymous 2hRP2pyrioNo ratings yet
- Critical Reviews in Oncology / HematologyDocument9 pagesCritical Reviews in Oncology / HematologyNadya MagfiraNo ratings yet
- Celulas Germinales CADocument9 pagesCelulas Germinales CAamenos18No ratings yet
- Neoplasia - Pathology SattarDocument3 pagesNeoplasia - Pathology SattarGursimranKaurSehgalNo ratings yet
- Uterine and Prostate CancerDocument13 pagesUterine and Prostate CancerSheana TmplNo ratings yet
- Anti-Mu Llerian Hormone: Ovarian Reserve Testing and Its Potential Clinical ImplicationsDocument14 pagesAnti-Mu Llerian Hormone: Ovarian Reserve Testing and Its Potential Clinical ImplicationsSri IriantiNo ratings yet
- Hum. Reprod. 2009 Scarpellini 2703 8Document6 pagesHum. Reprod. 2009 Scarpellini 2703 8MitarXNo ratings yet
- Gestational Trophoblastic NeoplasiaDocument18 pagesGestational Trophoblastic NeoplasiaTanuj sai kumarNo ratings yet
- MR 28 Agustus-1Document7 pagesMR 28 Agustus-1BramaNo ratings yet
- Pathogenesis of Testicular Germ Cell Tumours: Leendert H. J. Looijenga and J. Wolter OosterhuisDocument11 pagesPathogenesis of Testicular Germ Cell Tumours: Leendert H. J. Looijenga and J. Wolter OosterhuisLatansa DinaNo ratings yet
- Ovarian Malignant Germ Cell Tumors PDFDocument26 pagesOvarian Malignant Germ Cell Tumors PDFFernanda RiveraNo ratings yet
- An Ovarian Granulosa Cell Tumor in A Patient With Polycystic Ovarian Syndrome: A Case ReportDocument5 pagesAn Ovarian Granulosa Cell Tumor in A Patient With Polycystic Ovarian Syndrome: A Case ReportIJAR JOURNALNo ratings yet
- Potential Diagnostic Biomarkers For Human Uterine Mesenchymal Tumours Especially LMP2 1i and Cyclin E1 Differential ExpressionsDocument5 pagesPotential Diagnostic Biomarkers For Human Uterine Mesenchymal Tumours Especially LMP2 1i and Cyclin E1 Differential ExpressionsEditor IJTSRDNo ratings yet
- No Epitelial Ovarion Cancer 2018Document18 pagesNo Epitelial Ovarion Cancer 2018palomazul007No ratings yet
- IJSHR50Document12 pagesIJSHR50OMKAR DANDEKARNo ratings yet
- Hormone Replacement Therapy and Breast Cancer Lanc 12Document1 pageHormone Replacement Therapy and Breast Cancer Lanc 12KiloNo ratings yet
- POGS Practice Bulletin 3 FINALDocument6 pagesPOGS Practice Bulletin 3 FINALAileen TombocNo ratings yet
- Askep DICDocument11 pagesAskep DICInesya Zea RestartaNo ratings yet
- Sustain 2021 10652180Document6 pagesSustain 2021 10652180filonenko0448No ratings yet
- LI 1 AdenomyosisDocument4 pagesLI 1 AdenomyosisyasminNo ratings yet
- Non Epithelial Ovarian CancerDocument18 pagesNon Epithelial Ovarian CancerDR RISKA WAHYUNo ratings yet
- ESMO Testicular CancerDocument8 pagesESMO Testicular CancerBernadetta ViaNo ratings yet
- Organo TropismDocument12 pagesOrgano TropismKL TongsonNo ratings yet
- Micrornas and Recent Insights Into Pediatric Ovarian CancersDocument10 pagesMicrornas and Recent Insights Into Pediatric Ovarian CancersingeNo ratings yet
- Tog 12150Document10 pagesTog 12150saeed hasan saeedNo ratings yet
- LeiomyomaDocument35 pagesLeiomyomaChristian SalimNo ratings yet
- Leiomyoma 24Document35 pagesLeiomyoma 24Shannaz YudonoNo ratings yet
- Ferrets Oncology-MainDocument26 pagesFerrets Oncology-MainJOAQUINALONZOPEREIRANo ratings yet
- Gulbahce 2016Document5 pagesGulbahce 2016Valir HusleNo ratings yet
- 1 s2.0 S0944711307000578 MainDocument9 pages1 s2.0 S0944711307000578 MainMehboob AlamNo ratings yet
- Metaplastic CA 2 IhkDocument10 pagesMetaplastic CA 2 IhkWinta MayantiNo ratings yet
- Paper 4Document8 pagesPaper 4Miguel Lara RodriguezNo ratings yet
- Tmpe061 TMPDocument5 pagesTmpe061 TMPFrontiersNo ratings yet
- Clinical Endocrinology - 2022 - Coopmans - Molecular Genetic Testing in The Management of Pituitary DiseaseDocument12 pagesClinical Endocrinology - 2022 - Coopmans - Molecular Genetic Testing in The Management of Pituitary DiseasegarrobosNo ratings yet
- Clinical and Genetic Characterization of Basal Cell Carcinoma and Breast Cancer in A Single PatientDocument7 pagesClinical and Genetic Characterization of Basal Cell Carcinoma and Breast Cancer in A Single PatientMahmoud AlshahatNo ratings yet
- Acta 90 405Document6 pagesActa 90 405Eftychia GkikaNo ratings yet
- Theories of Endometrial CarcinogenesisDocument14 pagesTheories of Endometrial CarcinogenesisRisti SFNo ratings yet
- Novel KAT6B-KANSL1 Fusion Gene Identified by RNA Sequencing in Retroperitoneal Leiomyoma With T (10 17) (q22 q21)Document15 pagesNovel KAT6B-KANSL1 Fusion Gene Identified by RNA Sequencing in Retroperitoneal Leiomyoma With T (10 17) (q22 q21)Ioannis PanagopoulosNo ratings yet
- Endometrialcancer: Obesity, Genetics, and Targeted AgentsDocument17 pagesEndometrialcancer: Obesity, Genetics, and Targeted AgentsEftychia GkikaNo ratings yet
- Breast Cancer: An Endocrinologist's PerspectiveDocument34 pagesBreast Cancer: An Endocrinologist's PerspectiveSahirNo ratings yet
- Enf TrofoblasticaDocument10 pagesEnf TrofoblasticaDaniela CovarrubiasNo ratings yet
- Pineal Region Tumors PDFDocument15 pagesPineal Region Tumors PDFhimadriNo ratings yet
- PIIS0015028220303307Document8 pagesPIIS0015028220303307PabloNo ratings yet
- Mancari 2014Document6 pagesMancari 2014Berry BancinNo ratings yet
- Vogelstein Nm1087Document11 pagesVogelstein Nm1087Misha30No ratings yet
- Endometrial Cancer ESMO Clinical Practice GuidelinesDocument5 pagesEndometrial Cancer ESMO Clinical Practice Guidelinesjhon heriansyahNo ratings yet
- New WHO Classification of Pituitary Adenomas (4th Edition) : Assessment of Pituitary Transcription Factors and The Prognostic Histological FactorsDocument5 pagesNew WHO Classification of Pituitary Adenomas (4th Edition) : Assessment of Pituitary Transcription Factors and The Prognostic Histological FactorscristianescNo ratings yet
- Cancer de Cervix y EndometrioDocument24 pagesCancer de Cervix y EndometriomaritaradNo ratings yet
- Breast Disease: Diagnosis and Pathology, Volume 1From EverandBreast Disease: Diagnosis and Pathology, Volume 1Adnan AydinerNo ratings yet
- Multiple Myeloma and Other Plasma Cell NeoplasmsFrom EverandMultiple Myeloma and Other Plasma Cell NeoplasmsMeletios A. DimopoulosNo ratings yet
- Neuroendocrine Tumors: Surgical Evaluation and ManagementFrom EverandNeuroendocrine Tumors: Surgical Evaluation and ManagementJordan M. CloydNo ratings yet
- IDO1Document13 pagesIDO1Victor HaidarNo ratings yet
- Directed Expression of An Oncogene To Sertoli Cells in Transgenic Mice Using Mullerian Inhibiting Substance Regulatory SequencesDocument9 pagesDirected Expression of An Oncogene To Sertoli Cells in Transgenic Mice Using Mullerian Inhibiting Substance Regulatory SequencesVictor HaidarNo ratings yet
- Cancers: Role of Inflammation and Oxidative Stress Mediators in GliomasDocument20 pagesCancers: Role of Inflammation and Oxidative Stress Mediators in GliomasVictor HaidarNo ratings yet
- CYP11A1Document15 pagesCYP11A1Victor HaidarNo ratings yet
- Cardio Pulmonary ResuscitationDocument34 pagesCardio Pulmonary Resuscitationanimesh pandaNo ratings yet
- Pregnancy CycleDocument23 pagesPregnancy CycleJoel CGNo ratings yet
- Best Pediatric Neck MassesDocument116 pagesBest Pediatric Neck MassesOmar PeterNo ratings yet
- Root Canal Treatment: A Clinical Guide For Dental Students, General Dentists Who Like Doing Rcts and General Dentists Who Hate Doing RctsDocument10 pagesRoot Canal Treatment: A Clinical Guide For Dental Students, General Dentists Who Like Doing Rcts and General Dentists Who Hate Doing Rctskeerthipriya chatlaNo ratings yet
- ENG 10 PaperDocument5 pagesENG 10 PapervelascoaugustNo ratings yet
- Part 2 Evidence Evaluation PDFDocument8 pagesPart 2 Evidence Evaluation PDFNurul RafahNo ratings yet
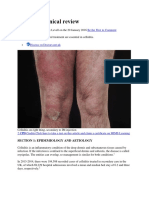
- Cellulitis: Clinical Review: Be The First To CommentDocument9 pagesCellulitis: Clinical Review: Be The First To CommentAnonymous 1nMTZWmzNo ratings yet
- Pat Physical Readiness Questionare 1Document1 pagePat Physical Readiness Questionare 1Rheez MelchorNo ratings yet
- AHS Subcutaneous TherapyDocument16 pagesAHS Subcutaneous TherapyTaylor NicholNo ratings yet
- LAS For Summative Assessment (Written Work Performance Task)Document2 pagesLAS For Summative Assessment (Written Work Performance Task)Angel Dela CruzNo ratings yet
- Travel Medicine: Health Quarantine 2021 Dr. Esther Meylina SipahutarDocument35 pagesTravel Medicine: Health Quarantine 2021 Dr. Esther Meylina SipahutarEsther Meyline XhypaNo ratings yet
- 301 ExperimentsWithAurasDocument5 pages301 ExperimentsWithAurasRaibaut PamiNo ratings yet
- 14 Application of Clinical Trial ResultsDocument5 pages14 Application of Clinical Trial ResultsDebanganaNo ratings yet
- Diploma in General Nursing PDFDocument7 pagesDiploma in General Nursing PDFTanwir HoussaynNo ratings yet
- Pediatric Hematology Oncology Ward Officer HandbookDocument40 pagesPediatric Hematology Oncology Ward Officer HandbookLetchumana KrishnanNo ratings yet
- Kalkaneal OsteotomiDocument11 pagesKalkaneal Osteotomitravma777No ratings yet
- Hypermagnesemia: From Wikipedia, The Free EncyclopediaDocument3 pagesHypermagnesemia: From Wikipedia, The Free EncyclopediayshenyNo ratings yet
- Spek Ven Avea Full ClioDocument3 pagesSpek Ven Avea Full ClioRinawatiNo ratings yet
- Makalah B.inggris RamaDocument8 pagesMakalah B.inggris RamaRama DianSyah PutraNo ratings yet
- Your Digestive SystemDocument2 pagesYour Digestive SystemTes Say EamNo ratings yet
- Infection Control: Dr.T.V.Rao MD Professor of MicrobiologyDocument41 pagesInfection Control: Dr.T.V.Rao MD Professor of Microbiologytummalapalli venkateswara rao100% (1)
- Assignment On "Fixed Partial Denture": Submitted byDocument5 pagesAssignment On "Fixed Partial Denture": Submitted byJeba Afia JebinNo ratings yet
- V6 Catalog GI - 230609Document8 pagesV6 Catalog GI - 230609bashir019No ratings yet
- Guía de La ACC Y AHA de 2020 para El Tratamiento de Pacientes Con Valvulopatías CardíacasDocument171 pagesGuía de La ACC Y AHA de 2020 para El Tratamiento de Pacientes Con Valvulopatías CardíacasMARIANA LINETH OROZCO RICARDONo ratings yet
- Benefits Manual: Wipro Mediclaim InsuranceDocument11 pagesBenefits Manual: Wipro Mediclaim InsuranceRavi TejaNo ratings yet
- Disseminated Tuberculosis in An AIDS/HIV-Infected Patient: AbstractDocument3 pagesDisseminated Tuberculosis in An AIDS/HIV-Infected Patient: AbstractAmelia Fitria DewiNo ratings yet