Download as pdf or txt
You might also like
- Brochure Cleaning General Global enDocument36 pagesBrochure Cleaning General Global ensetnjaprirodom80No ratings yet
- Bioethanol Production From Banana Peels: June 2016Document8 pagesBioethanol Production From Banana Peels: June 2016Charlie Micah AquinoNo ratings yet
- Biochemistry Lab Report: VNG - International UniversityDocument19 pagesBiochemistry Lab Report: VNG - International UniversityHà Anh Minh LêNo ratings yet
- WOR8294 Assaying and Refining of GoldDocument28 pagesWOR8294 Assaying and Refining of GoldRehan Sheikh100% (2)
- Accessing The Suitability of Using Banana Pith Juice As A Natural Coagulant For Textile Wastewater TreaDocument6 pagesAccessing The Suitability of Using Banana Pith Juice As A Natural Coagulant For Textile Wastewater TreaRizky PravitasariNo ratings yet
- Detoxification of Malachite Green and Textile Industrial Ef Uent by Penicillium OchrochloronDocument10 pagesDetoxification of Malachite Green and Textile Industrial Ef Uent by Penicillium OchrochloronJuan Diego CárdenasNo ratings yet
- Activated Sludge ProcessDocument48 pagesActivated Sludge Processrozma khanNo ratings yet
- Wastewater Treatment Using Cellulose Acetate Made From Rice HuskDocument8 pagesWastewater Treatment Using Cellulose Acetate Made From Rice HuskOmowu GbengaNo ratings yet
- Fruit Leather Inggris 5 DoneDocument3 pagesFruit Leather Inggris 5 DonefitriNo ratings yet
- Bioethanol Production From Alkaline-Pretreated Sugarcane BagasseDocument7 pagesBioethanol Production From Alkaline-Pretreated Sugarcane BagasseCuartos Amueblados JennyJohnnyNo ratings yet
- GSJ7016 (Aryan - Ramteke)Document20 pagesGSJ7016 (Aryan - Ramteke)CH24Aryan.RamtekeNo ratings yet
- Starch Research ProgressDocument15 pagesStarch Research ProgressEudkrenutNo ratings yet
- Enzymatic Digestibility of Pretreated Dewaxed Bamboo Residues As Feedstock For Bioethanol ProductionDocument7 pagesEnzymatic Digestibility of Pretreated Dewaxed Bamboo Residues As Feedstock For Bioethanol ProductionKevin Hernandez Duque100% (1)
- Mesocarpo de Coco Verde Pretratado Por Un Proceso Alcalino Como Materia Prima para La Producción de BioetanolDocument39 pagesMesocarpo de Coco Verde Pretratado Por Un Proceso Alcalino Como Materia Prima para La Producción de BioetanolNancy EspinosaNo ratings yet
- Natural Coagulant For Waste Water TreatmentDocument5 pagesNatural Coagulant For Waste Water TreatmentDhiyyah Mardhiyyah100% (1)
- Bioresource TechnologyDocument8 pagesBioresource TechnologyHerlin HerliansahNo ratings yet
- (PDF) Bioethanol Production From Banana PeelsDocument1 page(PDF) Bioethanol Production From Banana PeelsMidyaa TariqNo ratings yet
- Vegaandrade 2020Document11 pagesVegaandrade 2020VeronicaSandyMonzonTerronesNo ratings yet
- 7 PBDocument7 pages7 PBat6315763No ratings yet
- Bioethanols 2Document39 pagesBioethanols 2ahmad buysyNo ratings yet
- Bioresource TechnologyDocument7 pagesBioresource TechnologyAbiel Angel GarciaNo ratings yet
- Purification of Grey Water Using The Natural MethodDocument7 pagesPurification of Grey Water Using The Natural MethodMamta AgarwalNo ratings yet
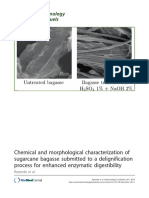
- Chemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityDocument19 pagesChemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityTabiita Huaman NorabuenaNo ratings yet
- Bioconversion of Posthydrolysed Autohydrolysis Liquors: An Alternative For Xylitol Production From Corn CobsDocument8 pagesBioconversion of Posthydrolysed Autohydrolysis Liquors: An Alternative For Xylitol Production From Corn CobsMaria Victoria Ortega RamosNo ratings yet
- AdsorptionDocument307 pagesAdsorptionΟδυσσεας ΚοψιδαςNo ratings yet
- Bioteknologi Pangan-Dina 2018Document9 pagesBioteknologi Pangan-Dina 2018achn_tyaNo ratings yet
- EnzymeDocument8 pagesEnzymeAlejandra LopezNo ratings yet
- Bioresources.: Bioconversion of Bamboo To Bioethanol Using The Two-Stage Organosolv and Alkali PretreatmentDocument9 pagesBioresources.: Bioconversion of Bamboo To Bioethanol Using The Two-Stage Organosolv and Alkali Pretreatmentcandra ndraNo ratings yet
- Oonkhanond 2017Document37 pagesOonkhanond 2017Gebby FebriliaNo ratings yet
- 2023 Efficient Extraction, Excellent Activity, and Microencapsulation of Flavonoids From Moringa Oleifera Leaves Extracted by Deep Eutectic SolventDocument15 pages2023 Efficient Extraction, Excellent Activity, and Microencapsulation of Flavonoids From Moringa Oleifera Leaves Extracted by Deep Eutectic SolventOlíviaGomesNo ratings yet
- Ghoshray 2015Document7 pagesGhoshray 2015ghoshraysreemoyeeNo ratings yet
- Biofuel PaperDocument6 pagesBiofuel PaperLy HangkheangNo ratings yet
- Flocullant Characteristics, Mechanism PolydadmacDocument9 pagesFlocullant Characteristics, Mechanism PolydadmaclalithvaradhanNo ratings yet
- Ijsrmjournal, Journal Manager, 20 IjsrmDocument10 pagesIjsrmjournal, Journal Manager, 20 IjsrmSheena Mae de LeonNo ratings yet
- Studies On Bio-Ethanol Production From Orange Peels Using Bacillus SubtilisDocument5 pagesStudies On Bio-Ethanol Production From Orange Peels Using Bacillus SubtilisAnoif Naputo AidnamNo ratings yet
- Sulbaran - 2020Document10 pagesSulbaran - 2020Belkis SulbaranNo ratings yet
- Elkady Et Al.. 2011. Bacillus Mojavensis Strain 32A, A Bioflocculant-Producing Bacterium Isolated From An Egyptian Salt Production PondDocument9 pagesElkady Et Al.. 2011. Bacillus Mojavensis Strain 32A, A Bioflocculant-Producing Bacterium Isolated From An Egyptian Salt Production PondAzb 711No ratings yet
- Sustainable Reclamation and Reuse of Industrial Wastewater Including Membrane Bioreactor TechnologiesDocument7 pagesSustainable Reclamation and Reuse of Industrial Wastewater Including Membrane Bioreactor TechnologiesadrianqNo ratings yet
- Comparative Study On Chemical Pretreatment Methods For Improving Enzymatic Digestibility of Crofton Weed StemDocument8 pagesComparative Study On Chemical Pretreatment Methods For Improving Enzymatic Digestibility of Crofton Weed StemLaura Aguilar NavasNo ratings yet
- V2i4 1160Document8 pagesV2i4 1160Ajay Kumar GantiNo ratings yet
- Polysaccharide PeelsDocument8 pagesPolysaccharide Peelsbouazizi souhirNo ratings yet
- Efecto de Pretratamientos en La Sacarificacion de Residuos de PiniaDocument8 pagesEfecto de Pretratamientos en La Sacarificacion de Residuos de PiniaEmma ValentinaNo ratings yet
- Dopado de AzufreDocument11 pagesDopado de AzufreChuy MartinezNo ratings yet
- Garbage Enzyme University SarawakDocument6 pagesGarbage Enzyme University SarawakSaints Burner ChristopherNo ratings yet
- Isolation and Screening of Hydrogen Producing Bacterial Strain From Sugarcane Bagasse Yard SoilDocument8 pagesIsolation and Screening of Hydrogen Producing Bacterial Strain From Sugarcane Bagasse Yard SoilATSNo ratings yet
- Bioresource Technology: Sinu Kumari, Debabrata DasDocument10 pagesBioresource Technology: Sinu Kumari, Debabrata DasMoha Abu-selmiahNo ratings yet
- Archive of SID: Removal of Surfactant From Industrial Wastewaters by Coagulation Flocculation ProcessDocument6 pagesArchive of SID: Removal of Surfactant From Industrial Wastewaters by Coagulation Flocculation ProcessrudymerchoNo ratings yet
- 10 1016@j Procbio 2014 10 012Document9 pages10 1016@j Procbio 2014 10 012LiriamNo ratings yet
- Potential Use of Eco-Enzyme For The Treatment of Metal Based EffluentDocument7 pagesPotential Use of Eco-Enzyme For The Treatment of Metal Based EffluentAdvennie NuhujananNo ratings yet
- Deep Eutectic Solvents for Efficient and Selective Extraction of α-Glucosidase Inhibitors from Waste Seeds of Refined Betel NutsDocument11 pagesDeep Eutectic Solvents for Efficient and Selective Extraction of α-Glucosidase Inhibitors from Waste Seeds of Refined Betel NutsCsaba AndrásNo ratings yet
- Role of Fermentation Technology in Sewage Waste Water Fatima DarakhshanDocument7 pagesRole of Fermentation Technology in Sewage Waste Water Fatima DarakhshanAreebah IftikharNo ratings yet
- Tongco2020 Article EnhancementOfHydrolysisAndBiogDocument9 pagesTongco2020 Article EnhancementOfHydrolysisAndBiogJim TsikasNo ratings yet
- Acid Hydrolysis of Sugarcane Bagasse For Lactic Acid ProductionDocument8 pagesAcid Hydrolysis of Sugarcane Bagasse For Lactic Acid ProductionAndressaGonçalvesNo ratings yet
- Fungal Biodegradation and Enzymatic Activities On Sawdust: March 2021Document10 pagesFungal Biodegradation and Enzymatic Activities On Sawdust: March 2021zmn zmnNo ratings yet
- Biomass Fractionation 3Document87 pagesBiomass Fractionation 3mackachytamysNo ratings yet
- tmp64AC TMPDocument8 pagestmp64AC TMPFrontiersNo ratings yet
- Removal of Organic Pollutants From Pharmaceutical Industry Wastewater by Chemical Coagulation and Bentonite Clay PretreatmentDocument4 pagesRemoval of Organic Pollutants From Pharmaceutical Industry Wastewater by Chemical Coagulation and Bentonite Clay PretreatmentRaluca RusuNo ratings yet
- Radhika 2012Document10 pagesRadhika 2012Pendi Adi MertaNo ratings yet
- Industrial Crops & Products: Felipe Fernandes-Klajn, Juan Miguel Romero-García, Manuel J. Díaz, Eulogio CastroDocument9 pagesIndustrial Crops & Products: Felipe Fernandes-Klajn, Juan Miguel Romero-García, Manuel J. Díaz, Eulogio Castrolucia789No ratings yet
- Accelerated Desalting and Purification of Silk Fibroin in A CaCl2-EtOH-H2O Ternary System by Excess Isopropanol ExtractionDocument17 pagesAccelerated Desalting and Purification of Silk Fibroin in A CaCl2-EtOH-H2O Ternary System by Excess Isopropanol ExtractionHozefa DhilaNo ratings yet
- Technology For Treatment of Lipid-Rich Wastewater and Pipelines Clogged by Lipids Using Bacterial PreparationDocument9 pagesTechnology For Treatment of Lipid-Rich Wastewater and Pipelines Clogged by Lipids Using Bacterial PreparationAlacránAlemánNo ratings yet
- Fungi and Lignocellulosic BiomassFrom EverandFungi and Lignocellulosic BiomassChristian P KubicekNo ratings yet
- Biological Sludge Minimization and Biomaterials/Bioenergy Recovery TechnologiesFrom EverandBiological Sludge Minimization and Biomaterials/Bioenergy Recovery TechnologiesEtienne PaulNo ratings yet
- My TestDocument8 pagesMy TestkrisnuNo ratings yet
- J Polymertesting 2008 09 006 PDFDocument8 pagesJ Polymertesting 2008 09 006 PDFShovon HalderNo ratings yet
- Exercise 9 (Acyl Compounds Soaps and Detergents)Document6 pagesExercise 9 (Acyl Compounds Soaps and Detergents)Johan Tadlas0% (1)
- Distance Learning Programme: Pre-Medical: Leader Test Series / Joint Package CourseDocument8 pagesDistance Learning Programme: Pre-Medical: Leader Test Series / Joint Package CourseGames MinixNo ratings yet
- Acids, Alkalis and Salts RevisionDocument19 pagesAcids, Alkalis and Salts RevisionJames EzardNo ratings yet
- Recent Developments in Multifunctional Coatings For Solar Panel Applications A Review PDFDocument28 pagesRecent Developments in Multifunctional Coatings For Solar Panel Applications A Review PDFkjjNo ratings yet
- Catalysts in Production of Biodiesel: A Review: K. Narasimharao, Adam Lee, and Karen WilsonDocument12 pagesCatalysts in Production of Biodiesel: A Review: K. Narasimharao, Adam Lee, and Karen WilsonFlorence IrhibhogbeNo ratings yet
- 0620 m17 QP 62 PDFDocument8 pages0620 m17 QP 62 PDFMinakshiNo ratings yet
- Basics Fundamentals of Liquid Chromatography HPLCDocument35 pagesBasics Fundamentals of Liquid Chromatography HPLCDr-Abdulsamie Hassan Alta'eeNo ratings yet
- Robin Murphy Nature's Materia Medica: LeseprobeDocument22 pagesRobin Murphy Nature's Materia Medica: LeseprobeBibhakar Dutta0% (1)
- Inkjet-Printed High Mobility Transparent-Oxide SemiconductorsDocument5 pagesInkjet-Printed High Mobility Transparent-Oxide Semiconductorsjulio perezNo ratings yet
- Descriptive Model of Laterite Type Bauxite DepositsDocument3 pagesDescriptive Model of Laterite Type Bauxite DepositsErsina Rakhma MoechsinNo ratings yet
- Bullock Et Al-2018-Geology TodayDocument5 pagesBullock Et Al-2018-Geology TodayABERA ERENANo ratings yet
- Personal Care Formulary - 2015Document28 pagesPersonal Care Formulary - 2015Evelina Kassabova100% (4)
- Chapter 2 Carbohydrates PDFDocument158 pagesChapter 2 Carbohydrates PDFAlyssum MarieNo ratings yet
- Module 4: Alkenes: Nomenclature, Preparation and ReactionDocument2 pagesModule 4: Alkenes: Nomenclature, Preparation and ReactionPeña, RodolfoNo ratings yet
- Solvent Free Sonochemical Namboodiri2002Document3 pagesSolvent Free Sonochemical Namboodiri2002Sherif Dei BukariNo ratings yet
- SolutionsDocument509 pagesSolutionssknoushad126No ratings yet
- Graphene Cement Concrete WhitePaper 2021Document30 pagesGraphene Cement Concrete WhitePaper 2021tytaityNo ratings yet
- 1 s2.0 S0956053X22000605 MainDocument11 pages1 s2.0 S0956053X22000605 MainIsraelPala-RosasNo ratings yet
- Chemistry 2017Document18 pagesChemistry 2017samar ahmedNo ratings yet
- Product Bulletin - PHPADocument1 pageProduct Bulletin - PHPAEliz MelizaNo ratings yet
- Grade 10 Science Unit 10 Eng.Document4 pagesGrade 10 Science Unit 10 Eng.Liza MathewNo ratings yet
- Isoelectric Focusing ElectrophoresisDocument11 pagesIsoelectric Focusing ElectrophoresisMadala IndraniNo ratings yet
- 2019-2020 Condensation PolymersDocument8 pages2019-2020 Condensation PolymersAgustina DenaNo ratings yet
- Practicals Final 10th Biology 1584103824Document19 pagesPracticals Final 10th Biology 1584103824nitinNo ratings yet
- 3.2.8. Sterile Single-Use Plastic Syringes-2Document2 pages3.2.8. Sterile Single-Use Plastic Syringes-2Avinash Kumar33% (3)