
Coordination of Morphogenesis and Cell-Fate
Coordination of Morphogenesis and Cell-Fate
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5822)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
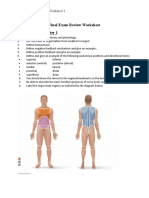
- Final Exam Review Worksheet Introduction-Chapter 1Document30 pagesFinal Exam Review Worksheet Introduction-Chapter 1Jensen BarrettNo ratings yet
- Principles of Fungicide Resistance: Fast FactsDocument5 pagesPrinciples of Fungicide Resistance: Fast FactsSOUMITA MITRANo ratings yet
- Estimation of Total Free Amino AcidDocument6 pagesEstimation of Total Free Amino AcidSOUMITA MITRANo ratings yet
- Effects of Uncouplers and Inhibitors of The Electron Transport-Oxidative Phosphorylation SystemDocument6 pagesEffects of Uncouplers and Inhibitors of The Electron Transport-Oxidative Phosphorylation SystemSOUMITA MITRANo ratings yet
- FungicideResistance 2Document60 pagesFungicideResistance 2SOUMITA MITRANo ratings yet
- CHN Ncm112b Rle RequirementDocument27 pagesCHN Ncm112b Rle RequirementPatricia VasquezNo ratings yet
- Bones of The Axial SkeletonDocument4 pagesBones of The Axial SkeletonJasminKate SutacioNo ratings yet
- BIOLOGY NOTES For StuffDocument42 pagesBIOLOGY NOTES For StuffTudorNedelcuNo ratings yet
- Paper: Sophie Cavallero, Diane Riccobono, Michel Drouet, and Sabine FrançoisDocument16 pagesPaper: Sophie Cavallero, Diane Riccobono, Michel Drouet, and Sabine FrançoisMalik AzeemNo ratings yet
- Grade 9 Lesson1Document60 pagesGrade 9 Lesson1Julius LitaNo ratings yet
- Yoga For Arthritis The Complete GuideDocument465 pagesYoga For Arthritis The Complete GuideRomina Mascetti100% (2)
- A New View of Pain As A Homeostatic EmotionDocument5 pagesA New View of Pain As A Homeostatic EmotionJulian GorositoNo ratings yet
- Healing Energy - Part 1 Historical BackgroundDocument6 pagesHealing Energy - Part 1 Historical BackgroundDianaNo ratings yet
- Branches of Zoology: EthologyDocument2 pagesBranches of Zoology: EthologymuhammadismailNo ratings yet
- Neonatal AnaesthesiaDocument30 pagesNeonatal Anaesthesiavenkatesh chowdaryNo ratings yet
- AnthelminticsDocument8 pagesAnthelminticsshaitabliganNo ratings yet
- 99 Words Every High School Graduate Should KnowDocument3 pages99 Words Every High School Graduate Should KnowMila Tania TjNo ratings yet
- Educ1 2.1 TopicDocument31 pagesEduc1 2.1 TopicChriane AldamarNo ratings yet
- Introductory Lecture 3rd Year MBBSDocument12 pagesIntroductory Lecture 3rd Year MBBSPakeezaNo ratings yet
- Endocrinology and CriminologyDocument224 pagesEndocrinology and CriminologyRob OvalleNo ratings yet
- Environment International: Ryoiti Kiyama, Yuko Wada-KiyamaDocument30 pagesEnvironment International: Ryoiti Kiyama, Yuko Wada-KiyamaFABIANA PASSAMANINo ratings yet
- Sudan Medical Specialization Board 2Document37 pagesSudan Medical Specialization Board 2Dania Zaid100% (1)
- Activity 1: Can You Justify Me?: Hormone Yes No ReasonDocument2 pagesActivity 1: Can You Justify Me?: Hormone Yes No ReasonMa Rijubelyn Macaranas CapinpinNo ratings yet
- Urine Routine Examination: U804173 Id No MDocument1 pageUrine Routine Examination: U804173 Id No MTB AhmedNo ratings yet
- 3656 8873 1 SMDocument10 pages3656 8873 1 SMDino KaninoNo ratings yet
- Lab Rep 5Document27 pagesLab Rep 5Reign RosadaNo ratings yet
- ReportViewer3 CBCDocument1 pageReportViewer3 CBCMuhammad Munawar BuzdarNo ratings yet
- Lab ResultDocument1 pageLab ResultTanvir HamimNo ratings yet
- Exercise Standards For Testing and TrainingDocument62 pagesExercise Standards For Testing and TrainingRelviGuzmanApazaNo ratings yet
- NCM 112 Activity 3A Surgical Instruments: Mayo StraightDocument9 pagesNCM 112 Activity 3A Surgical Instruments: Mayo StraightJustine Simeon lagunzadNo ratings yet
- Handbook Canine Feline NutritionDocument332 pagesHandbook Canine Feline Nutritiontayna ritielle100% (1)
- ImmunoSeroLec M2Document10 pagesImmunoSeroLec M2ela kikayNo ratings yet
- AquacultureDocument8 pagesAquacultureWahyu W'cas PamungkasNo ratings yet
- Enzymes Organic ChemistryDocument6 pagesEnzymes Organic Chemistryمحمد الكبيسيNo ratings yet










































































Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5822)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Final Exam Review Worksheet Introduction-Chapter 1Document30 pagesFinal Exam Review Worksheet Introduction-Chapter 1Jensen BarrettNo ratings yet
- Principles of Fungicide Resistance: Fast FactsDocument5 pagesPrinciples of Fungicide Resistance: Fast FactsSOUMITA MITRANo ratings yet
- Estimation of Total Free Amino AcidDocument6 pagesEstimation of Total Free Amino AcidSOUMITA MITRANo ratings yet
- Effects of Uncouplers and Inhibitors of The Electron Transport-Oxidative Phosphorylation SystemDocument6 pagesEffects of Uncouplers and Inhibitors of The Electron Transport-Oxidative Phosphorylation SystemSOUMITA MITRANo ratings yet
- FungicideResistance 2Document60 pagesFungicideResistance 2SOUMITA MITRANo ratings yet
- CHN Ncm112b Rle RequirementDocument27 pagesCHN Ncm112b Rle RequirementPatricia VasquezNo ratings yet
- Bones of The Axial SkeletonDocument4 pagesBones of The Axial SkeletonJasminKate SutacioNo ratings yet
- BIOLOGY NOTES For StuffDocument42 pagesBIOLOGY NOTES For StuffTudorNedelcuNo ratings yet
- Paper: Sophie Cavallero, Diane Riccobono, Michel Drouet, and Sabine FrançoisDocument16 pagesPaper: Sophie Cavallero, Diane Riccobono, Michel Drouet, and Sabine FrançoisMalik AzeemNo ratings yet
- Grade 9 Lesson1Document60 pagesGrade 9 Lesson1Julius LitaNo ratings yet
- Yoga For Arthritis The Complete GuideDocument465 pagesYoga For Arthritis The Complete GuideRomina Mascetti100% (2)
- A New View of Pain As A Homeostatic EmotionDocument5 pagesA New View of Pain As A Homeostatic EmotionJulian GorositoNo ratings yet
- Healing Energy - Part 1 Historical BackgroundDocument6 pagesHealing Energy - Part 1 Historical BackgroundDianaNo ratings yet
- Branches of Zoology: EthologyDocument2 pagesBranches of Zoology: EthologymuhammadismailNo ratings yet
- Neonatal AnaesthesiaDocument30 pagesNeonatal Anaesthesiavenkatesh chowdaryNo ratings yet
- AnthelminticsDocument8 pagesAnthelminticsshaitabliganNo ratings yet
- 99 Words Every High School Graduate Should KnowDocument3 pages99 Words Every High School Graduate Should KnowMila Tania TjNo ratings yet
- Educ1 2.1 TopicDocument31 pagesEduc1 2.1 TopicChriane AldamarNo ratings yet
- Introductory Lecture 3rd Year MBBSDocument12 pagesIntroductory Lecture 3rd Year MBBSPakeezaNo ratings yet
- Endocrinology and CriminologyDocument224 pagesEndocrinology and CriminologyRob OvalleNo ratings yet
- Environment International: Ryoiti Kiyama, Yuko Wada-KiyamaDocument30 pagesEnvironment International: Ryoiti Kiyama, Yuko Wada-KiyamaFABIANA PASSAMANINo ratings yet
- Sudan Medical Specialization Board 2Document37 pagesSudan Medical Specialization Board 2Dania Zaid100% (1)
- Activity 1: Can You Justify Me?: Hormone Yes No ReasonDocument2 pagesActivity 1: Can You Justify Me?: Hormone Yes No ReasonMa Rijubelyn Macaranas CapinpinNo ratings yet
- Urine Routine Examination: U804173 Id No MDocument1 pageUrine Routine Examination: U804173 Id No MTB AhmedNo ratings yet
- 3656 8873 1 SMDocument10 pages3656 8873 1 SMDino KaninoNo ratings yet
- Lab Rep 5Document27 pagesLab Rep 5Reign RosadaNo ratings yet
- ReportViewer3 CBCDocument1 pageReportViewer3 CBCMuhammad Munawar BuzdarNo ratings yet
- Lab ResultDocument1 pageLab ResultTanvir HamimNo ratings yet
- Exercise Standards For Testing and TrainingDocument62 pagesExercise Standards For Testing and TrainingRelviGuzmanApazaNo ratings yet
- NCM 112 Activity 3A Surgical Instruments: Mayo StraightDocument9 pagesNCM 112 Activity 3A Surgical Instruments: Mayo StraightJustine Simeon lagunzadNo ratings yet
- Handbook Canine Feline NutritionDocument332 pagesHandbook Canine Feline Nutritiontayna ritielle100% (1)
- ImmunoSeroLec M2Document10 pagesImmunoSeroLec M2ela kikayNo ratings yet
- AquacultureDocument8 pagesAquacultureWahyu W'cas PamungkasNo ratings yet
- Enzymes Organic ChemistryDocument6 pagesEnzymes Organic Chemistryمحمد الكبيسيNo ratings yet