Download as pdf or txt
You might also like
- Cycle SyncingDocument43 pagesCycle SyncingHeShot MeDown100% (1)
- RMU Test ReportDocument3 pagesRMU Test ReportGANESH K75% (4)
- Airport Literature StudyDocument15 pagesAirport Literature StudySoundar Rajan100% (2)
- Fate MapsDocument25 pagesFate MapsImran MohsinNo ratings yet
- GastrulationDocument1 pageGastrulationClaire BolalinNo ratings yet
- Bio ProjectDocument17 pagesBio ProjectRiya Jaiswal100% (1)
- Sex-Determination System - WikipediaDocument61 pagesSex-Determination System - WikipediaDeepak maliNo ratings yet
- Genetic Basis of InheritanceDocument14 pagesGenetic Basis of InheritanceaditiNo ratings yet
- Economic ZoologyDocument47 pagesEconomic ZoologyAntony SebastianNo ratings yet
- Growth & Devlopment in Insurance SectorDocument36 pagesGrowth & Devlopment in Insurance SectorVijay90% (29)
- Unit: 08 Placentation in Mammals: Developmental Biology Cr. Hours: 4 (3+1)Document30 pagesUnit: 08 Placentation in Mammals: Developmental Biology Cr. Hours: 4 (3+1)Task BirdNo ratings yet
- Endoplasmic ReticulumDocument36 pagesEndoplasmic ReticulumShikhaj ArmaanNo ratings yet
- 3 - Physiology and Anatomy of ReproductionDocument13 pages3 - Physiology and Anatomy of ReproductionjuhNo ratings yet
- Sericulture - FormattedDocument18 pagesSericulture - FormattedllldNo ratings yet
- B.Sc. (Ug) Zoology Syllabus: Tumkur University TumakuruDocument40 pagesB.Sc. (Ug) Zoology Syllabus: Tumkur University TumakuruPraveen NekkantiNo ratings yet
- Sperm Morphology - 2019 FinalDocument44 pagesSperm Morphology - 2019 FinalDr Jitu moni DasNo ratings yet
- Soil Analysis and Agricultural SurveyDocument38 pagesSoil Analysis and Agricultural SurveySarvesh RautNo ratings yet
- SpirogyraDocument25 pagesSpirogyraAmir HamzaNo ratings yet
- Chick NeurulationDocument14 pagesChick NeurulationSophia RubiaNo ratings yet
- Biomolecules Chemistry AssignmentDocument27 pagesBiomolecules Chemistry AssignmentUvan VijayNo ratings yet
- StainigDocument38 pagesStainigyuppie_raj2175No ratings yet
- Human Reroduction by Digambar Mali.Document22 pagesHuman Reroduction by Digambar Mali.Surge E-Solution Pvt.Ltd.100% (1)
- Cryopreservation: Anuja Kamath, M.SC Bangalore Assisted Conception CentreDocument50 pagesCryopreservation: Anuja Kamath, M.SC Bangalore Assisted Conception CentreAl MamunNo ratings yet
- Surrogacyin India Ethicaland Legal AspectDocument8 pagesSurrogacyin India Ethicaland Legal AspectannnoyynnmussNo ratings yet
- Fate Maps: Course: B.Sc. (H) Zoology VI Semester Paper: Developmental Biology Faculty: Dr. Priya GoelDocument17 pagesFate Maps: Course: B.Sc. (H) Zoology VI Semester Paper: Developmental Biology Faculty: Dr. Priya GoelSusmita PalNo ratings yet
- Application of BiotechnologyDocument7 pagesApplication of BiotechnologyJaideep DasNo ratings yet
- Database ConceptsDocument31 pagesDatabase ConceptswafasaNo ratings yet
- Cell JunctionsDocument56 pagesCell JunctionsDrGagandeep Singh100% (3)
- Haemocytometer Cell CountsDocument2 pagesHaemocytometer Cell CountsSnigdha GuptaNo ratings yet
- Developmental Biology, 12th Edition (Michael J.F. Barresi, Scott F. Gilbert)Document48 pagesDevelopmental Biology, 12th Edition (Michael J.F. Barresi, Scott F. Gilbert)Nita DelinaNo ratings yet
- Gene Regulation in Prokaryotes 11022017Document55 pagesGene Regulation in Prokaryotes 11022017Nur Syamimi0% (1)
- Microsporogenesis Callose Deposition and Its SignificanceDocument10 pagesMicrosporogenesis Callose Deposition and Its SignificanceGYANRANJAN MAHALIKNo ratings yet
- 40 Life Cycle HerdmaniaDocument7 pages40 Life Cycle HerdmaniaAyushiNo ratings yet
- Neurulation Refers To The Folding Process in Vertebrate Embryos, Which Includes TheDocument6 pagesNeurulation Refers To The Folding Process in Vertebrate Embryos, Which Includes Theaaliya saaheenNo ratings yet
- Somatic Embryogenesis: Meaning, History, Principles, Protocols and ImportanceDocument11 pagesSomatic Embryogenesis: Meaning, History, Principles, Protocols and ImportanceAlka PriyadarshiniNo ratings yet
- Project Report ON Mushroom Cultivation Technology (ELM-422) : Submitted byDocument63 pagesProject Report ON Mushroom Cultivation Technology (ELM-422) : Submitted byRobin SidhuNo ratings yet
- Payroll FullDocument117 pagesPayroll FullushaNo ratings yet
- Fish Parasite Heavy Mental and Gene ExpressionDocument22 pagesFish Parasite Heavy Mental and Gene ExpressionJackyLeeNo ratings yet
- 01 - Embryology (Theory)Document54 pages01 - Embryology (Theory)Simran Kashyap100% (1)
- MSC BotanyDocument102 pagesMSC BotanydragonwarriorNo ratings yet
- Report On The Visit To National Dairy Research InstituteDocument15 pagesReport On The Visit To National Dairy Research InstitutemuhsinNo ratings yet
- Stem Cells and Organ CultureDocument18 pagesStem Cells and Organ CultureGEETA MOHANNo ratings yet
- Fetal MembranesDocument17 pagesFetal MembranesQaiser InayatNo ratings yet
- Laboratory Safety and RegulationsLaboratory SkillsDocument4 pagesLaboratory Safety and RegulationsLaboratory SkillsIklilKamarulZamanNo ratings yet
- Cleavage, Blastula, Gastrula, NeurulaDocument41 pagesCleavage, Blastula, Gastrula, NeurulaHyenaNo ratings yet
- The Cell Cycle & Cell DivisionDocument30 pagesThe Cell Cycle & Cell DivisionamitNo ratings yet
- The Morphology of The Post-Gastric Alimentary CanalDocument18 pagesThe Morphology of The Post-Gastric Alimentary CanalAnca MihalcescuNo ratings yet
- Usage of Cell PhonesDocument35 pagesUsage of Cell PhonesJayapradha SiduNo ratings yet
- 1 - Sex DeterminationDocument18 pages1 - Sex Determinationmis_administratorNo ratings yet
- The Cell CycleDocument72 pagesThe Cell CycleJerry Jeroum RegudoNo ratings yet
- Dokumen - Tips - Biology Investigatory Project Class 12Document17 pagesDokumen - Tips - Biology Investigatory Project Class 12Aryan PatelNo ratings yet
- Automatic Water Pump Controller PDFDocument44 pagesAutomatic Water Pump Controller PDFmphatsoNo ratings yet
- Development Developmental Biology EmbryologyDocument6 pagesDevelopment Developmental Biology EmbryologyBiju ThomasNo ratings yet
- Honours Syllabus PDFDocument48 pagesHonours Syllabus PDFPANKAJ MondalNo ratings yet
- Biology Project On Drug Resistance in BacteriaDocument26 pagesBiology Project On Drug Resistance in Bacteriaashisham014No ratings yet
- Early Development in BirdsDocument53 pagesEarly Development in BirdsMuqaddas NadeemNo ratings yet
- Endoplasmic ReticulumDocument30 pagesEndoplasmic ReticulumMuhammad Junaid Iqbal100% (1)
- Protochordata-Characters & PhylogenyDocument4 pagesProtochordata-Characters & PhylogenyAakash VNo ratings yet
- Development of The KidneyDocument62 pagesDevelopment of The KidneyRIUNGU PENINAH GATWIRINo ratings yet
- Plant Taxonomy NotesDocument38 pagesPlant Taxonomy Notesopolla nianorNo ratings yet
- EmbryologyDocument12 pagesEmbryologyKara KaneNo ratings yet
- Developmental Biology of Frog: SpermDocument21 pagesDevelopmental Biology of Frog: SpermRavindra MadurNo ratings yet
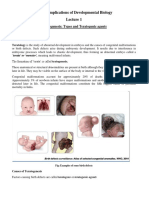
- Unit 5: Implications of Developmental Biology: Teratogenesis: Types and Teratogenic AgentsDocument12 pagesUnit 5: Implications of Developmental Biology: Teratogenesis: Types and Teratogenic AgentsAmar Kant JhaNo ratings yet
- Stem Cells - Sources, Characteristics, Types, Uses - Developmental Biology - Microbe NotesDocument5 pagesStem Cells - Sources, Characteristics, Types, Uses - Developmental Biology - Microbe NotesAmar Kant JhaNo ratings yet
- Developing BalstulationDocument2 pagesDeveloping BalstulationAmar Kant JhaNo ratings yet
- Cell LineageDocument7 pagesCell LineageAmar Kant JhaNo ratings yet
- Spider-81 Hardware Spec 7.7Document7 pagesSpider-81 Hardware Spec 7.7KonradNo ratings yet
- Sri Lank An Airline IndustryDocument29 pagesSri Lank An Airline IndustryTuan RifkhanNo ratings yet
- 11 Chemistry Module 2Document19 pages11 Chemistry Module 2SpongeBob SquarePants Fidget ToysNo ratings yet
- TinyMobileRobots TinySurveyor FlyerDocument4 pagesTinyMobileRobots TinySurveyor FlyerfelipeNo ratings yet
- SONTU Flat Panel Dectector CatalogDocument6 pagesSONTU Flat Panel Dectector CatalogWajahat SheikhNo ratings yet
- Form 67 Delivery Unloading and Loading of Materials and PlantDocument6 pagesForm 67 Delivery Unloading and Loading of Materials and PlantMohamed MedhioubNo ratings yet
- TapingDocument3 pagesTapingEdeza MatarumNo ratings yet
- Sewing Skills Checklist PDFDocument44 pagesSewing Skills Checklist PDFyemkem100% (1)
- NEW Sales Tax Invoice - 2023-07-27T164634.549Document1 pageNEW Sales Tax Invoice - 2023-07-27T164634.549Saadat IrfanNo ratings yet
- HwyconstDocument23 pagesHwyconstAmulie JarjuseyNo ratings yet
- Uremic EncephalophatyDocument48 pagesUremic EncephalophatySindi LadayaNo ratings yet
- HEI Tech Sheet 110Document15 pagesHEI Tech Sheet 110Suganya LokeshNo ratings yet
- Allotrope Partners Indonesia Renewable Energy Project AssociateDocument2 pagesAllotrope Partners Indonesia Renewable Energy Project AssociateRizal ZulkarnaenNo ratings yet
- Straumann Product Catalogue 2018 2019Document260 pagesStraumann Product Catalogue 2018 2019kllasikalleNo ratings yet
- Law of Mother Earth BoliviaDocument3 pagesLaw of Mother Earth Boliviarahul banerjeeNo ratings yet
- The Normal DistributionDocument30 pagesThe Normal DistributionJohn Rich CaidicNo ratings yet
- TPS54160 1.5-A, 60-V, Step-Down DC/DC Converter With Eco-Mode™Document57 pagesTPS54160 1.5-A, 60-V, Step-Down DC/DC Converter With Eco-Mode™sbrhomeNo ratings yet
- Annual Report 2014 PDFDocument153 pagesAnnual Report 2014 PDFகோகுல் இராNo ratings yet
- Jack and The Beanstalk Treatment-2Document10 pagesJack and The Beanstalk Treatment-2api-668257195No ratings yet
- 9701 s02 ErDocument14 pages9701 s02 ErHubbak KhanNo ratings yet
- Quarter 3 - Las No. 1 (Week 5-7) Active Recreation (Street and Hip-Hop Dances) (Pe10Pf-Iiia-H-39, Pe10Pf-Lllc-H-45)Document11 pagesQuarter 3 - Las No. 1 (Week 5-7) Active Recreation (Street and Hip-Hop Dances) (Pe10Pf-Iiia-H-39, Pe10Pf-Lllc-H-45)hakkensNo ratings yet
- HPB21-0457 FINAL Submittal 09-07-2021 - Ritz Sagamore LOIDocument13 pagesHPB21-0457 FINAL Submittal 09-07-2021 - Ritz Sagamore LOIthe next miamiNo ratings yet
- AbseilingDocument12 pagesAbseilingMurah Rezeki Cikgu WafiNo ratings yet
- Kohima Nagaland LBDocument1 pageKohima Nagaland LBIndia TreadingNo ratings yet
- 9 Exercise Physiology Handout 2011Document20 pages9 Exercise Physiology Handout 2011maraj687No ratings yet
- Process SequenceDocument2 pagesProcess SequenceUmesh SakhareliyaNo ratings yet
- Petroleum Engineering Lab 1 (PGE313) Experiment 2 Determination of PorosityDocument13 pagesPetroleum Engineering Lab 1 (PGE313) Experiment 2 Determination of PorosityAli AbubakarNo ratings yet