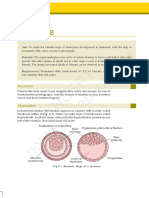
Download as pdf or txt
You might also like
- This Study Resource Was: Drosophila Simulation - Patterns of HeredityDocument8 pagesThis Study Resource Was: Drosophila Simulation - Patterns of HeredityFUN PHYSICS FOR NEET67% (3)
- CSIWildlife StudentWS1 AnsKeyDocument4 pagesCSIWildlife StudentWS1 AnsKeyIshan Sharma44% (18)
- AP Biology Lab Seven: Genetics of OrganismsDocument8 pagesAP Biology Lab Seven: Genetics of OrganismsCoolAsianDude67% (15)
- Hydrotaxis and Phototaxis of Terrestrial Isopods Armadillidium VulgareDocument7 pagesHydrotaxis and Phototaxis of Terrestrial Isopods Armadillidium VulgareCynthia GriffithNo ratings yet
- Drosophila Lab ReportDocument7 pagesDrosophila Lab ReportTim HepburnNo ratings yet
- ALV - ALV Utility ProgramDocument4 pagesALV - ALV Utility ProgramSubash AbapNo ratings yet
- Automotive Safety HandbookDocument2 pagesAutomotive Safety HandbookSenthil Muthu Kumar T0% (1)
- Research Paper - Phototaxis in Zophobas MorioDocument9 pagesResearch Paper - Phototaxis in Zophobas Morioapi-393048315No ratings yet
- Advantages of Having A Lateralized BrainDocument3 pagesAdvantages of Having A Lateralized BrainJohn S DoeNo ratings yet
- Reconocimiento Conspecífico en Palomas (Columba Livia) Utilizando Imágenes de Video DinámicasDocument11 pagesReconocimiento Conspecífico en Palomas (Columba Livia) Utilizando Imágenes de Video DinámicasMi-liz RamirezNo ratings yet
- TMP 2663Document9 pagesTMP 2663FrontiersNo ratings yet
- What's The Deal With Birds?: Scientific Journal of Research and ReviewsDocument4 pagesWhat's The Deal With Birds?: Scientific Journal of Research and ReviewsResul AYDINNo ratings yet
- Learning by Association in PlantsDocument9 pagesLearning by Association in PlantsmishelNo ratings yet
- Drosophilia Manuscript 1Document6 pagesDrosophilia Manuscript 1api-638364487No ratings yet
- Learning and Cognition in InsectsDocument13 pagesLearning and Cognition in InsectsmarianaNo ratings yet
- Lubow, Moore (1959)Document5 pagesLubow, Moore (1959)Dagon EscarabajoNo ratings yet
- 3 Anderson 2001Document5 pages3 Anderson 2001claudia osorioNo ratings yet
- 24 Angelaki Nature 2004Document5 pages24 Angelaki Nature 2004sebNo ratings yet
- Drosophila Lab Write UpDocument13 pagesDrosophila Lab Write Upapi-585837255No ratings yet
- Functional Ecology - 2021 - Franklin - Cracks in The Mirror Hypothesis High Specularity Does Not Reduce Detection orDocument10 pagesFunctional Ecology - 2021 - Franklin - Cracks in The Mirror Hypothesis High Specularity Does Not Reduce Detection orDinh Tan TranNo ratings yet
- Agee y Patterson 1983 Trampas Stomoxys HaematobiaDocument6 pagesAgee y Patterson 1983 Trampas Stomoxys HaematobiajuanNo ratings yet
- © Ncert Not To Be Republished: Exercise 8Document5 pages© Ncert Not To Be Republished: Exercise 8Ankit YadavNo ratings yet
- 2005-Effect of Different IlluminationDocument6 pages2005-Effect of Different IlluminationberilNo ratings yet
- Life&BheaviourDocument12 pagesLife&BheaviourGraziana RussoNo ratings yet
- Monkeys, Mirrors, Mark Tests and Minds: Steven M. Platek and Sarah L. LevinDocument2 pagesMonkeys, Mirrors, Mark Tests and Minds: Steven M. Platek and Sarah L. LevinSenad R. AvdovićNo ratings yet
- Atypical Eye Gaze Perception inDocument21 pagesAtypical Eye Gaze Perception inLeandro FerrazNo ratings yet
- 2011 Ryabov, Polyakov, Bibikov - Characterization of Beam Patterns of Bottlenose Dolphin in The Transverse PlaneDocument6 pages2011 Ryabov, Polyakov, Bibikov - Characterization of Beam Patterns of Bottlenose Dolphin in The Transverse PlanebvromanovNo ratings yet
- Introduction To The Science of EcologyDocument14 pagesIntroduction To The Science of Ecologyt.a.mojtahedNo ratings yet
- Laboratory Assessment 3Document4 pagesLaboratory Assessment 3Juliano, Jhanielle Faye B.No ratings yet
- Sexual Dichromatism in The Blue Fornted Amazon (Amazona Aestiva) Revealed by Multiple Angel SpectrometryDocument8 pagesSexual Dichromatism in The Blue Fornted Amazon (Amazona Aestiva) Revealed by Multiple Angel SpectrometryDiego LeónNo ratings yet
- House Fly Oviposition Inhibition by Larvae of Hermetia Illucens, The Black Soldier FlyDocument7 pagesHouse Fly Oviposition Inhibition by Larvae of Hermetia Illucens, The Black Soldier FlySantos RojoNo ratings yet
- Symmetry: When and Why Did Brains Break Symmetry?Document14 pagesSymmetry: When and Why Did Brains Break Symmetry?Anonymous UjdVtuTtqNo ratings yet
- Lab Activity 2Document8 pagesLab Activity 2Mellya RizkiNo ratings yet
- Kunst-Wilson (1980) Affective Discrimination of Stimuli That Cannot Be RecognizedDocument2 pagesKunst-Wilson (1980) Affective Discrimination of Stimuli That Cannot Be RecognizedJohn TownsendNo ratings yet
- Valenzuela SDocument6 pagesValenzuela SVaisGaNo ratings yet
- Invertebrate Welfare Where Is The Real Evidence For Conscious Affective StatesDocument2 pagesInvertebrate Welfare Where Is The Real Evidence For Conscious Affective StatesGergő GáspárNo ratings yet
- Learning With Half A Brain: David D. Lent, Marianna Pinte R, Nicholas J. StrausfeldDocument12 pagesLearning With Half A Brain: David D. Lent, Marianna Pinte R, Nicholas J. StrausfeldAbhishekParmarNo ratings yet
- Articulo CristianDocument5 pagesArticulo CristianDaBid Lopez RodriguezNo ratings yet
- Genetics of Mouse Behavior: Interactions With Laboratory EnvironmentDocument3 pagesGenetics of Mouse Behavior: Interactions With Laboratory EnvironmentAndrea Castro HaroNo ratings yet
- Imitative Learning in Male Japanese Quail (Coturnix Japonica) Using The Two-Action MethodDocument5 pagesImitative Learning in Male Japanese Quail (Coturnix Japonica) Using The Two-Action MethodWahyu TedjaNo ratings yet
- Social Modulation of Learning in Rats: ResearchDocument9 pagesSocial Modulation of Learning in Rats: ResearchSammer BurgosNo ratings yet
- Postoperative Environmental Enrichment Attenuates Fimbria-Fornix Lesion-Induced Impairments in Morris Maze PerformanceDocument8 pagesPostoperative Environmental Enrichment Attenuates Fimbria-Fornix Lesion-Induced Impairments in Morris Maze PerformanceJean Pierre Chastre LuzaNo ratings yet
- American Association For The Advancement of ScienceDocument3 pagesAmerican Association For The Advancement of SciencePriss SaezNo ratings yet
- Jurnal Sishew AmphibiDocument7 pagesJurnal Sishew AmphibishafaNo ratings yet
- The Use of Computed Tomography in AvianDocument7 pagesThe Use of Computed Tomography in AvianDaniel BernalNo ratings yet
- Behavioural ProcessesDocument11 pagesBehavioural ProcessesharisankarhsNo ratings yet
- Article 7Document7 pagesArticle 7hammmamira08No ratings yet
- Altenhoff2019 Protocol InferringOrthologyAndParalogyDocument28 pagesAltenhoff2019 Protocol InferringOrthologyAndParalogyCalvince OdhiamboNo ratings yet
- All Rodents Are Not The Same: A Modern Synthesis of Cortical OrganizationDocument44 pagesAll Rodents Are Not The Same: A Modern Synthesis of Cortical Organizationmarej312No ratings yet
- 1 s2.0 S0168159196010933 MainDocument17 pages1 s2.0 S0168159196010933 MainLucy BrittainNo ratings yet
- 1 s2.0 0031938494900221 MainDocument5 pages1 s2.0 0031938494900221 MainjonathonmarinisNo ratings yet
- Affective Discrimination of The Stimuli That Cannot Be RecognizedDocument3 pagesAffective Discrimination of The Stimuli That Cannot Be Recognizedbarrylyndon1967No ratings yet
- Fenton, M (2012)Document11 pagesFenton, M (2012)lina maaria lopezNo ratings yet
- CSIWildlife Worksheet1Document5 pagesCSIWildlife Worksheet1Noah HolznerNo ratings yet
- Elcoro 2008Document8 pagesElcoro 2008Moko NugrohoNo ratings yet
- Behavior Differentiation Between Wild Quail and Domestic QuailDocument6 pagesBehavior Differentiation Between Wild Quail and Domestic QuailCamila AndreaNo ratings yet
- Vision Research: Frances J. Rucker, Josh WallmanDocument9 pagesVision Research: Frances J. Rucker, Josh Wallmans397665300No ratings yet
- Determining Acceptance of The 9:3:3:1 Ratio in Fruit Fly Crosses Using The Chi Squared TestDocument7 pagesDetermining Acceptance of The 9:3:3:1 Ratio in Fruit Fly Crosses Using The Chi Squared TestFUN PHYSICS FOR NEETNo ratings yet
- Linkage Disequilibrium in The Human GenomeDocument7 pagesLinkage Disequilibrium in The Human GenomedioufNo ratings yet
- 1 s2.0 S0014299903012780 MainDocument17 pages1 s2.0 S0014299903012780 MainOscar René Hernández PérezNo ratings yet
- Morphological Variation in a Population of the Snake, Tantilla gracilis Baird and GirardFrom EverandMorphological Variation in a Population of the Snake, Tantilla gracilis Baird and GirardNo ratings yet
- Baseline Values of Immunologic Parameters in The LDocument7 pagesBaseline Values of Immunologic Parameters in The LSofía DalmazzoNo ratings yet
- Lord 1956Document21 pagesLord 1956Sofía DalmazzoNo ratings yet
- The Scaling of Eye Size With Body Mass in Birds:, Doi: 10.1098/rspb.1999.0652Document9 pagesThe Scaling of Eye Size With Body Mass in Birds:, Doi: 10.1098/rspb.1999.0652Sofía DalmazzoNo ratings yet
- Flight Distance and Eye Size in Birds: EthologyDocument8 pagesFlight Distance and Eye Size in Birds: EthologySofía DalmazzoNo ratings yet
- Chap 1-Data and Information Assignment1 With AnswerDocument2 pagesChap 1-Data and Information Assignment1 With AnswerPoornima CNNo ratings yet
- USHANATESANDocument15 pagesUSHANATESANSaravanan KrNo ratings yet
- TOPIC 1: Different Nautical ChartsDocument9 pagesTOPIC 1: Different Nautical ChartsRaymund Suaybaguio100% (2)
- Internship ReportDocument11 pagesInternship Reportkefyalew TNo ratings yet
- Nota SKAA4412-Lab PDFDocument32 pagesNota SKAA4412-Lab PDFsyamsul anwarNo ratings yet
- Master Thesis PDFDocument4 pagesMaster Thesis PDFafcngxbbx100% (2)
- Assessment of Aleatory and Model Uncertainties For Non-LinearDocument131 pagesAssessment of Aleatory and Model Uncertainties For Non-LinearGenNo ratings yet
- Lecture Ii-5: The Bose-Fermi Correspondence and Its Applications Edward Witten 5.1. 2-Dimensional Gauge Theories With FermionsDocument9 pagesLecture Ii-5: The Bose-Fermi Correspondence and Its Applications Edward Witten 5.1. 2-Dimensional Gauge Theories With FermionsluisdanielNo ratings yet
- Design ParametersDocument6 pagesDesign Parameterswasylika877745No ratings yet
- Datasheet PlateCatcher 2018Document4 pagesDatasheet PlateCatcher 2018Secure LifeNo ratings yet
- PreviewpdfDocument57 pagesPreviewpdframiru da maeztuNo ratings yet
- IWRBS 3 - WHLP - Week 4Document2 pagesIWRBS 3 - WHLP - Week 4jolinamarizNo ratings yet
- Cleaning Measurews in OtDocument4 pagesCleaning Measurews in OttanishaNo ratings yet
- User InterfacesDocument1 pageUser InterfacesВо ВаNo ratings yet
- A Study On Distributed Leadership Practices and Its Impact On Teaching and LearningDocument18 pagesA Study On Distributed Leadership Practices and Its Impact On Teaching and LearningGlobal Research and Development ServicesNo ratings yet
- Fingpt: Open-Source Financial Large Language ModelsDocument7 pagesFingpt: Open-Source Financial Large Language ModelsMaks GutNo ratings yet
- Arte Util - Stalker - Osservatorio NomadeDocument1 pageArte Util - Stalker - Osservatorio NomadeNicoleNo ratings yet
- Fundamentals of Coal Mine Roof SupportDocument21 pagesFundamentals of Coal Mine Roof Supportindira intanNo ratings yet
- PHILOSOPHYDocument2 pagesPHILOSOPHYgandarraNo ratings yet
- Nebosh How To Pass Your Open Book Exam On The First AttemptDocument4 pagesNebosh How To Pass Your Open Book Exam On The First Attemptdp.cavozziNo ratings yet
- Lesson Plan Template 2021Document3 pagesLesson Plan Template 2021api-550991282No ratings yet
- Portfolio Activity 4Document5 pagesPortfolio Activity 4Joel Christian MascariñaNo ratings yet
- 3.18-3.21- Trần Thị Trà MyDocument6 pages3.18-3.21- Trần Thị Trà MyThị Trà My TrầnNo ratings yet
- Rubic Project 2 - Video AnimationDocument1 pageRubic Project 2 - Video AnimationAliana RosuriNo ratings yet
- D300s Advanced - Shooting Custom Menu Banks 12-09-8pDocument8 pagesD300s Advanced - Shooting Custom Menu Banks 12-09-8ptelmopNo ratings yet
- Design Heuristic WorkshopDocument22 pagesDesign Heuristic WorkshopMaryel LaraNo ratings yet
- Observation in CarDocument8 pagesObservation in CarNanda FujianaNo ratings yet
- New PepeortDocument25 pagesNew PepeortUsama Arfat MalikNo ratings yet