Download as pdf or txt
You might also like
- Karps Cell and Molecular Biology Ninth 9Th Edition Gerald Karp Full ChapterDocument67 pagesKarps Cell and Molecular Biology Ninth 9Th Edition Gerald Karp Full Chapterronald.dancy562100% (10)
- Textbook First Aid For The Basic Sciences General Principles Tao Le Ebook All Chapter PDFDocument53 pagesTextbook First Aid For The Basic Sciences General Principles Tao Le Ebook All Chapter PDFshane.roscoe731100% (15)
- Ebook PDF Understanding Biology 3rd Edition by Kenneth Mason PDFDocument41 pagesEbook PDF Understanding Biology 3rd Edition by Kenneth Mason PDFrichard.rogers744100% (40)
- Kumpulan Quiz CourseraDocument13 pagesKumpulan Quiz CourseraNadya Azzahra Salsabilla SaragihNo ratings yet
- HRW Água Hidrogenada Melhora No Fígado de Ratos EnvenenadosDocument6 pagesHRW Água Hidrogenada Melhora No Fígado de Ratos EnvenenadosNai ValenteNo ratings yet
- Structural Characterization and Antioxidant Activities of A NovelDocument10 pagesStructural Characterization and Antioxidant Activities of A Novelela.sofiaNo ratings yet
- Said AhmedDocument19 pagesSaid Ahmedahmed mahdiNo ratings yet
- Wang 2016 - Reviews On Mechanisms of in Vitro Antioxidant Activity of PolysaccharidesDocument14 pagesWang 2016 - Reviews On Mechanisms of in Vitro Antioxidant Activity of PolysaccharidesDaniellyNo ratings yet
- Antioxidant Status and Oxidative Stress Markers of White Faeces Syndrome-Infected Pacific White Shrimp (Litopenaeus Vannamei Boone)Document15 pagesAntioxidant Status and Oxidative Stress Markers of White Faeces Syndrome-Infected Pacific White Shrimp (Litopenaeus Vannamei Boone)Igor BaltaNo ratings yet
- Molecules 27 07756Document13 pagesMolecules 27 07756Sayuri hernandez vargasNo ratings yet
- Redox-Sensitive Irinotecan Liposomes With Active Ultra-High Loading andDocument9 pagesRedox-Sensitive Irinotecan Liposomes With Active Ultra-High Loading andwu79513No ratings yet
- The Impact of Plasma ActivatedWater Treatment On The Phenolic Profile, Vitamins Content, Antioxidant and Enzymatic Activities of Rocket-Salad LeavesDocument15 pagesThe Impact of Plasma ActivatedWater Treatment On The Phenolic Profile, Vitamins Content, Antioxidant and Enzymatic Activities of Rocket-Salad LeavesAHMED MOHAMMED MAHMOUD MUSTAFANo ratings yet
- Marinedrugs 14 00203Document16 pagesMarinedrugs 14 00203FABIO ESTEBAN HERRERA ROCHANo ratings yet
- 1 s2.0 S0032579121004648 MainDocument9 pages1 s2.0 S0032579121004648 MainVeterinary nijamati lok sewaNo ratings yet
- Antioxidant Capability and Efficacy of MDocument8 pagesAntioxidant Capability and Efficacy of MalexanderjfernandesNo ratings yet
- Fig 1Document6 pagesFig 1조민성No ratings yet
- Cui (2022)Document9 pagesCui (2022)Ally BNo ratings yet
- Chiết xuất lá atisô, như chất ức chế AKR1B1, làm giảm mức sorbitol trong thấu kính mắt chuột trong điều kiện glucose cao ex vivoDocument7 pagesChiết xuất lá atisô, như chất ức chế AKR1B1, làm giảm mức sorbitol trong thấu kính mắt chuột trong điều kiện glucose cao ex vivoLại Ngọc MaiNo ratings yet
- Baranowska 2020Document14 pagesBaranowska 2020Vanja TodorovocNo ratings yet
- Ameliorated Effect of Lactobacillus Plantarum Scs2 On The Oxidative Stress in Hepg2 Cells Induced by Afb1Document9 pagesAmeliorated Effect of Lactobacillus Plantarum Scs2 On The Oxidative Stress in Hepg2 Cells Induced by Afb1Yeni Katon SujarnokoNo ratings yet
- Structural Characterization and Antioxidant Activity of Alkali-Extracted Polysaccharides FroDocument10 pagesStructural Characterization and Antioxidant Activity of Alkali-Extracted Polysaccharides FroMich SanzNo ratings yet
- Purification of Polysaccharides From Cordyceps Militaris and Their Anti Hypoxic EffectDocument6 pagesPurification of Polysaccharides From Cordyceps Militaris and Their Anti Hypoxic EffectRepositorioNo ratings yet
- Lycopene Inhibits LPS-Induced Proinflammatory Mediator Inducible Nitric Oxide Synthase in Mouse Macrophage CellsDocument6 pagesLycopene Inhibits LPS-Induced Proinflammatory Mediator Inducible Nitric Oxide Synthase in Mouse Macrophage CellsNguyen Thanh SangNo ratings yet
- JFNR 7 9 5Document4 pagesJFNR 7 9 5edoNo ratings yet
- Therapeutic Potential and Biological Applications of-PMCDocument26 pagesTherapeutic Potential and Biological Applications of-PMCHarendra ChunduruNo ratings yet
- Agaricus Bisporus Fucogalactan Structural Characterization and Pharmacological ApproachesDocument8 pagesAgaricus Bisporus Fucogalactan Structural Characterization and Pharmacological ApproachesHuy NguyenNo ratings yet
- In Vitro Antioxidant Activity of SilymarinDocument12 pagesIn Vitro Antioxidant Activity of Silymarinbane95nsNo ratings yet
- Abstract F MauretteDocument1 pageAbstract F MauretteWanderley OliveiraNo ratings yet
- Rajakumari Et Al2020Document32 pagesRajakumari Et Al2020Gabriel GursenNo ratings yet
- Molecules: in Vitro Antioxidant Activities of Three Red Wine PolyphenolsDocument13 pagesMolecules: in Vitro Antioxidant Activities of Three Red Wine PolyphenolslolaNo ratings yet
- Life 2273833Document30 pagesLife 2273833a.drioicheNo ratings yet
- Sharath 05 (Biological Macromolecules)Document10 pagesSharath 05 (Biological Macromolecules)Dr. Sharath Kumar M NNo ratings yet
- Cahova 2014Document12 pagesCahova 2014yasmin matterNo ratings yet
- Articulo Curvas Estandar PDFDocument7 pagesArticulo Curvas Estandar PDFJorge GutierrezNo ratings yet
- Foods 12 03314Document14 pagesFoods 12 03314ela.sofiaNo ratings yet
- JMB026-07-07 Fdoc 1Document8 pagesJMB026-07-07 Fdoc 1Samantha Garcia NovoaNo ratings yet
- Sustainability 11 02933Document15 pagesSustainability 11 02933Mohammad ShofieNo ratings yet
- Molecules 27 03432abtsDocument17 pagesMolecules 27 03432abtsgtenienteNo ratings yet
- Trace Elements Analysis by Icpoes After Microwave Digestion of M 2014Document5 pagesTrace Elements Analysis by Icpoes After Microwave Digestion of M 2014Aytekin GaribliNo ratings yet
- Nae Et Al 2008Document7 pagesNae Et Al 2008BeatriceMihaelaRaduNo ratings yet
- Effect of Sodium Benzoate On Liver and Kidney Lipid Peroxidation and Antioxidant Enzymes in MiceDocument7 pagesEffect of Sodium Benzoate On Liver and Kidney Lipid Peroxidation and Antioxidant Enzymes in MiceDiana PașanNo ratings yet
- Characterization and Immunological Activity of Exopolysaccharide From Lacticaseibacillus Paracasei GL1 Isolated From Tibetan Kefir GrainsDocument18 pagesCharacterization and Immunological Activity of Exopolysaccharide From Lacticaseibacillus Paracasei GL1 Isolated From Tibetan Kefir GrainsOlga JakšićNo ratings yet
- Odbayar 2006Document5 pagesOdbayar 2006Corona FingerNo ratings yet
- Toxicity of Monocrotophos in Freshwater Bivalve, Lamellidens Marginalis, Using Different MarkersDocument9 pagesToxicity of Monocrotophos in Freshwater Bivalve, Lamellidens Marginalis, Using Different MarkersIJRASETPublicationsNo ratings yet
- Accepted Manuscript: DeserticolaDocument21 pagesAccepted Manuscript: DeserticolaFUCKMENo ratings yet
- 10jafc58 5380Document7 pages10jafc58 5380mailNo ratings yet
- Liu 2014Document10 pagesLiu 2014RicoNo ratings yet
- 10.1007 - s11694 017 9573 7Document10 pages10.1007 - s11694 017 9573 7Juan MartínezNo ratings yet
- Anti Cancer Mechanism and Possibility ofDocument4 pagesAnti Cancer Mechanism and Possibility ofOlivia Pebri RahayuNo ratings yet
- FT IR and FT Raman Fingerprints of Flavonoids - A Review - 2022 - Food ChemistryDocument9 pagesFT IR and FT Raman Fingerprints of Flavonoids - A Review - 2022 - Food ChemistryvanbanbinhdinhNo ratings yet
- Biomolecules 12 00444Document12 pagesBiomolecules 12 00444Madiha AshrafNo ratings yet
- Toxic Effects of Fungicide Sheathmar To Worm, Tubifex TubifexDocument12 pagesToxic Effects of Fungicide Sheathmar To Worm, Tubifex TubifexIJRASETPublicationsNo ratings yet
- A Moderate-Fat Diet With One Avocado Per Day Increases Plasma Antioxidants and Decreases The Oxidation of Small, Dense LDL in Adults With Overweight and Obesity RCT 2019Document9 pagesA Moderate-Fat Diet With One Avocado Per Day Increases Plasma Antioxidants and Decreases The Oxidation of Small, Dense LDL in Adults With Overweight and Obesity RCT 2019Kelly DickinsonNo ratings yet
- Molecular Mechanism of Formalin-Induced Toxicity and Its ManagementDocument8 pagesMolecular Mechanism of Formalin-Induced Toxicity and Its ManagementbiayuniarNo ratings yet
- Shifa AisyahDocument9 pagesShifa AisyahIndahNo ratings yet
- Dobrowolska-Iwanek Et-Al Hplc-Dad Method For The Quantitative Determination 2020Document7 pagesDobrowolska-Iwanek Et-Al Hplc-Dad Method For The Quantitative Determination 2020Dayanne ZeladaNo ratings yet
- Efect of Priobotics On Growth Profiles and Hematology 0f Nile TilapiaDocument7 pagesEfect of Priobotics On Growth Profiles and Hematology 0f Nile TilapiaYousaf JamalNo ratings yet
- Jurnal 1Document9 pagesJurnal 1Yunita Carolline PanggabeanNo ratings yet
- Sigma SaponinDocument3 pagesSigma Saponinkate2494No ratings yet
- Journal of Biodiversity and Environmental Sciences (JBES)Document9 pagesJournal of Biodiversity and Environmental Sciences (JBES)International Network For Natural SciencesNo ratings yet
- Foods 09 01033 v2Document15 pagesFoods 09 01033 v2nguyendongthucNo ratings yet
- Faheem ArticleDocument15 pagesFaheem ArticleFaheem UllahNo ratings yet
- Quercetin Inhibits Advanced Glycation End Product Formation by Trapping Methylglyoxal and GlyoxalDocument8 pagesQuercetin Inhibits Advanced Glycation End Product Formation by Trapping Methylglyoxal and GlyoxalYeny ChristianaNo ratings yet
- S BI Plants Hormones 09 TDocument12 pagesS BI Plants Hormones 09 TparthakerNo ratings yet
- General Pharmacology: Pharmacodynamics: Dr. Sahil Kumar NautiyalDocument58 pagesGeneral Pharmacology: Pharmacodynamics: Dr. Sahil Kumar NautiyalYashasv BhatnagarNo ratings yet
- Drug Targets Lecture Notes 2Document15 pagesDrug Targets Lecture Notes 2sriNo ratings yet
- Pathophysiology of AsthmaDocument71 pagesPathophysiology of AsthmaChin ChanNo ratings yet
- Cytokines BluesDocument15 pagesCytokines BluesantonioNo ratings yet
- Project Report Ias-Nasi-Insa Summer Research Fellowship Programme 2010Document30 pagesProject Report Ias-Nasi-Insa Summer Research Fellowship Programme 2010Bisesh PadhiNo ratings yet
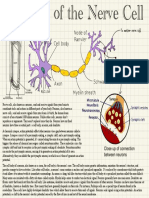
- Biology PosterDocument1 pageBiology PosterNoobmaster666No ratings yet
- 7.013 Pset 4 PDFDocument12 pages7.013 Pset 4 PDF15klaNo ratings yet
- Receptor PhysiologyDocument34 pagesReceptor PhysiologyKarl Torres Uganiza RmtNo ratings yet
- Hospital and Clinical PharmacyDocument18 pagesHospital and Clinical PharmacySayeeda MohammedNo ratings yet
- Test Bank For Basic Immunology 3rd Edition Abul K AbbasDocument7 pagesTest Bank For Basic Immunology 3rd Edition Abul K AbbasAdam Seger100% (42)
- Practice MCQ SynNeuroDocument4 pagesPractice MCQ SynNeuroMawj Mandwie100% (1)
- Abstract. Hair Regression and Balding Are Distressing ConcernsDocument7 pagesAbstract. Hair Regression and Balding Are Distressing ConcernsAPilgrimGodNo ratings yet
- PathwaysDocument9 pagesPathwaysIvan John ClementNo ratings yet
- Problem Set 2 EndocrinologyDocument12 pagesProblem Set 2 EndocrinologyJessica EtieneNo ratings yet
- AP Biology SupplementDocument86 pagesAP Biology SupplementLama DiriyehNo ratings yet
- Intracellular Trafficking and SortingDocument67 pagesIntracellular Trafficking and SortingSanthoshsv 143No ratings yet
- Partoza, J WS Cel - MolDocument6 pagesPartoza, J WS Cel - MolJenny PartozaNo ratings yet
- Chapter 2, Unit 1, Human Anatomy and Physiology 1, B Pharmacy 1st Sem, Carewell PharmaDocument21 pagesChapter 2, Unit 1, Human Anatomy and Physiology 1, B Pharmacy 1st Sem, Carewell Pharmaom441693No ratings yet
- Biochem - Hormone and Signal TransductionDocument36 pagesBiochem - Hormone and Signal TransductionPrincess Jeyan PagatpatanNo ratings yet
- Tissue EngineeringDocument17 pagesTissue EngineeringAntonio LoureiroNo ratings yet
- Chapter 2 Subcellular Organization of The Nervous Sys 2014 From MoleculesDocument30 pagesChapter 2 Subcellular Organization of The Nervous Sys 2014 From MoleculesJaisson Jahziel Blancas GarciaNo ratings yet
- Do WN Lo Ad Ed Fro M: Regulation of Airway Smooth Muscle Force Generation by Integrin-Containing Adhesion ComplexesDocument3 pagesDo WN Lo Ad Ed Fro M: Regulation of Airway Smooth Muscle Force Generation by Integrin-Containing Adhesion ComplexesIrmaBummielfNo ratings yet
- IrdaDocument763 pagesIrdaSalamatus riskiyahNo ratings yet
- Biomarkers-Emerging Applications of Biomarkers in HealthcareDocument3 pagesBiomarkers-Emerging Applications of Biomarkers in Healthcaresulabh chandraNo ratings yet
- NHT - Exs. 1-38.8.31.16Document175 pagesNHT - Exs. 1-38.8.31.16Spreter & PetiprinNo ratings yet