Download as doc, pdf, or txt
You might also like
- Immunology Exam Q's With AnswersDocument26 pagesImmunology Exam Q's With AnswersHavardKrovel88% (56)
- Janeways Immunobiology 9Th Edition Murphy Test Bank Full Chapter PDFDocument44 pagesJaneways Immunobiology 9Th Edition Murphy Test Bank Full Chapter PDFJamesWellsoeic100% (12)
- Oral Lichen Planus - An Update On Pathogenesis and TreatmentDocument12 pagesOral Lichen Planus - An Update On Pathogenesis and TreatmentserieschorkiNo ratings yet
- Jurnal 6Document28 pagesJurnal 6aprildrgNo ratings yet
- Oral Lichen Planus: An Update On Pathogenesis and Treatment: Address For CorrespondenceDocument6 pagesOral Lichen Planus: An Update On Pathogenesis and Treatment: Address For Correspondenceputriayu varasantiNo ratings yet
- N Lavanya P Jayanthi Umadevi K Rao K Ranganathan: Correspondence AddressDocument9 pagesN Lavanya P Jayanthi Umadevi K Rao K Ranganathan: Correspondence AddressAnnisa Hanif MetandaNo ratings yet
- Oral Lichen Planus An Update On Pathogenesis and TreatmentDocument21 pagesOral Lichen Planus An Update On Pathogenesis and TreatmentsevattapillaiNo ratings yet
- Pathogenesis of Oral Lichen Planus - A ReviewDocument7 pagesPathogenesis of Oral Lichen Planus - A ReviewintanNo ratings yet
- Jurnal 1Document5 pagesJurnal 1Dzaenap UlfahNo ratings yet
- Review Article: Oral Lichen Planus As A Preneoplastic Inflammatory ModelDocument9 pagesReview Article: Oral Lichen Planus As A Preneoplastic Inflammatory ModelFakhrur RaziNo ratings yet
- 32Document21 pages32gamhaelNo ratings yet
- DR Meenakshi MDS Oral PathologyDocument29 pagesDR Meenakshi MDS Oral PathologyDr. Meenakshi SinghalNo ratings yet
- Liqeun Plano PDFDocument8 pagesLiqeun Plano PDFjuan pablo meneses rojasNo ratings yet
- Tzanck SmearDocument5 pagesTzanck SmearLince WijoyoNo ratings yet
- Cytokine: E.S. Robinson, V.P. WerthDocument9 pagesCytokine: E.S. Robinson, V.P. WerthArthurAkkNo ratings yet
- Pathophysiology of Cutaneous Lupus Erythematosus: Review Open AccessDocument10 pagesPathophysiology of Cutaneous Lupus Erythematosus: Review Open AccesspdahlianaNo ratings yet
- Oral Lichen Planus: Clinical Presentation and Management: Paul C. Edwards, BSC, MSC, Dds Robert Kelsch, DMDDocument6 pagesOral Lichen Planus: Clinical Presentation and Management: Paul C. Edwards, BSC, MSC, Dds Robert Kelsch, DMDPieter JohnyNo ratings yet
- Lichen PlanusDocument109 pagesLichen Planuschgarg199No ratings yet
- Immunology of Periodontal DiseasesDocument9 pagesImmunology of Periodontal DiseasesoladunniNo ratings yet
- Arduino 2017Document8 pagesArduino 2017neetika guptaNo ratings yet
- OLPandupdate Carrozzo ThorpeDocument13 pagesOLPandupdate Carrozzo ThorpeAstrid Dwi SattiNo ratings yet
- Australian Dental Journal - 2010 - Schifter - Oral Mucosal Diseases The Inflammatory DermatosesDocument16 pagesAustralian Dental Journal - 2010 - Schifter - Oral Mucosal Diseases The Inflammatory Dermatosesneetika guptaNo ratings yet
- The WHO-EORTC ClassificationDocument17 pagesThe WHO-EORTC ClassificationLagavulin2No ratings yet
- Matute SDocument5 pagesMatute SLUCASNo ratings yet
- White Lesions of The Oral MucosaDocument213 pagesWhite Lesions of The Oral MucosaJunaid Ramzan100% (1)
- Premalignant LesionsDocument76 pagesPremalignant LesionsGowri RaguNo ratings yet
- Premalignant Oral LesionsDocument94 pagesPremalignant Oral LesionssdinyantiNo ratings yet
- SepsisDocument2 pagesSepsisNannai02No ratings yet
- Fine-Needle Aspiration of Subcutaneous Panniculitis-Like T-Cell LymphomaDocument2 pagesFine-Needle Aspiration of Subcutaneous Panniculitis-Like T-Cell LymphomaMarta MudarraNo ratings yet
- Atypical Cellular Disorders: Kenneth L. Mcclain, Yasodha Natkunam, and Steven H. SwerdlowDocument14 pagesAtypical Cellular Disorders: Kenneth L. Mcclain, Yasodha Natkunam, and Steven H. SwerdlowhemendreNo ratings yet
- PAEDS 1a - 04.10.19 ThrombocytopeniaDocument15 pagesPAEDS 1a - 04.10.19 Thrombocytopenialotp12No ratings yet
- The Complement System and Human Autoimmune DiseasesDocument25 pagesThe Complement System and Human Autoimmune DiseasesCsr ArsNo ratings yet
- Pathology Lecture 2nd CourseDocument128 pagesPathology Lecture 2nd CourseAbdullah EssaNo ratings yet
- Chapter 53:: Paraneoplastic Pemphigus:: Grant J. Anhalt & Daniel MimouniDocument15 pagesChapter 53:: Paraneoplastic Pemphigus:: Grant J. Anhalt & Daniel MimouniFiraNo ratings yet
- Jced 7 E656Document4 pagesJced 7 E656María José VázquezNo ratings yet
- Lecture 03Document30 pagesLecture 03bsmls-f18-017No ratings yet
- WBC DisordersDocument17 pagesWBC DisordersAnaJovanovskaNo ratings yet
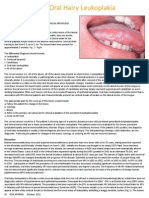
- Hairy LeukoplakiaDocument2 pagesHairy LeukoplakiaEka Septariana PuspaNo ratings yet
- HaematologyDocument25 pagesHaematologyMenziPhiwokuhleSukatiNo ratings yet
- Dermatomyositis Case7Document15 pagesDermatomyositis Case7Hanina Macataman MetmugNo ratings yet
- Publishers Take NoteaxicvDocument1 pagePublishers Take Noteaxicvwoolonion5No ratings yet
- Oral Lichen Planus An Enigma With Limited Treatment OptionsDocument4 pagesOral Lichen Planus An Enigma With Limited Treatment OptionsInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- SLEDocument42 pagesSLETrishenth FonsekaNo ratings yet
- Oral Lichen Planus and Lichenoid Reactions Etiopathogenesis Diagnosis, Management and Malignant TransformationDocument2 pagesOral Lichen Planus and Lichenoid Reactions Etiopathogenesis Diagnosis, Management and Malignant TransformationRosnah Binti ZainNo ratings yet
- Oral Lichen Planus PaperDocument17 pagesOral Lichen Planus PaperFakhrur RaziNo ratings yet
- Malignant Lymphoepithelial Lesion of The Parotid GlandDocument3 pagesMalignant Lymphoepithelial Lesion of The Parotid GlandJeffrey WibowoNo ratings yet
- Oral Lupus ErythematosusDocument9 pagesOral Lupus ErythematosusputriNo ratings yet
- EN + EosinofiliaDocument3 pagesEN + EosinofiliaAna Flavia BaptistaNo ratings yet
- Classic Versus Type II Enteropathy-Associated T-Cell LymphomaDocument5 pagesClassic Versus Type II Enteropathy-Associated T-Cell Lymphomaderek lauNo ratings yet
- 2006 Gosche Vick SemPedSurgeryDocument8 pages2006 Gosche Vick SemPedSurgeryResya I. NoerNo ratings yet
- Staphylococcal Skin InfectionsDocument25 pagesStaphylococcal Skin InfectionsteenaNo ratings yet
- Anatomy: Hem Atolog YDocument94 pagesAnatomy: Hem Atolog YaamirNo ratings yet
- Ocular Allergy: Abdulrahman Al MuammarDocument106 pagesOcular Allergy: Abdulrahman Al MuammarSugumar YathavanNo ratings yet
- All About VcamDocument8 pagesAll About VcamIkhsan AliNo ratings yet
- LP 160610182350Document25 pagesLP 160610182350Karthik TNo ratings yet
- Eye ImmunologyDocument35 pagesEye ImmunologySingo DemurNo ratings yet
- Shan Shal 2012Document15 pagesShan Shal 2012Jose AbadiaNo ratings yet
- Chronic Lymphocytic LeukemiaFrom EverandChronic Lymphocytic LeukemiaMichael HallekNo ratings yet
- Diagnosis of Cutaneous Lymphoid Infiltrates: A Visual Approach to Differential Diagnosis and Knowledge GapsFrom EverandDiagnosis of Cutaneous Lymphoid Infiltrates: A Visual Approach to Differential Diagnosis and Knowledge GapsNo ratings yet
- Autoimmune Diseases of Oral CavityDocument52 pagesAutoimmune Diseases of Oral Cavitylakshmi k sNo ratings yet
- Giant Cell LesionsDocument33 pagesGiant Cell Lesionslakshmi k sNo ratings yet
- 8 Haematoxylin and EosinDocument19 pages8 Haematoxylin and Eosinlakshmi k sNo ratings yet
- AmelogemnesisDocument114 pagesAmelogemnesislakshmi k sNo ratings yet
- StainingDocument81 pagesStaininglakshmi k sNo ratings yet
- S K Gupta, Binita Goswami Immunology For MBBS Arya Publishing CompanyDocument128 pagesS K Gupta, Binita Goswami Immunology For MBBS Arya Publishing Companyy6qqnhdvf2No ratings yet
- IMMUNOSERO (Quizlet)Document19 pagesIMMUNOSERO (Quizlet)Allyssa AniNo ratings yet
- 5a. Mekanisme Imunitas Seluler Dan HumoralDocument32 pages5a. Mekanisme Imunitas Seluler Dan HumoralGaluh Apsari SyafranNo ratings yet
- Immunology MCQDocument31 pagesImmunology MCQArulkumar Mani55% (11)
- Immunogen, Antigen, Epitope, HaptenDocument28 pagesImmunogen, Antigen, Epitope, HaptenIwan SutriswantoNo ratings yet
- Extracellular Matrix ScafoldDocument16 pagesExtracellular Matrix ScafoldmanishNo ratings yet
- Sistem Imun 2Document20 pagesSistem Imun 2CameliaMasrijalNo ratings yet
- UT Dallas Syllabus For Biol4345.001 06s Taught by John Burr (Burr)Document1 pageUT Dallas Syllabus For Biol4345.001 06s Taught by John Burr (Burr)UT Dallas Provost's Technology GroupNo ratings yet
- Histology BloodDocument6 pagesHistology BloodNeneng Cutin GanzonNo ratings yet
- Section 14 CELL-DYN 4000 Immuno T-Cell (CD3/4/8) Assay: 9140350A-October 2001Document58 pagesSection 14 CELL-DYN 4000 Immuno T-Cell (CD3/4/8) Assay: 9140350A-October 2001freedNo ratings yet
- PDF Middletons Allergy Essentials 1E 1 Har PSC Edition Robyn E Ohehir Fracp PHD Frcpath Ebook Full ChapterDocument63 pagesPDF Middletons Allergy Essentials 1E 1 Har PSC Edition Robyn E Ohehir Fracp PHD Frcpath Ebook Full Chapterkathy.cargle605100% (4)
- Immunomodulatory - Effect - of - Various - Anti 20170531 28954 Odcyzi With Cover Page v2Document14 pagesImmunomodulatory - Effect - of - Various - Anti 20170531 28954 Odcyzi With Cover Page v2Alex AlexNo ratings yet
- Material Suplementario Art. 5 PDFDocument29 pagesMaterial Suplementario Art. 5 PDFmaria alejandra parada aguilarNo ratings yet
- Samson Notes ALL IN ONEDocument855 pagesSamson Notes ALL IN ONEdentistmai24No ratings yet
- Can Res-2018-HPV16-T-cell - ELISpotDocument13 pagesCan Res-2018-HPV16-T-cell - ELISpotAlberto Rivas CidNo ratings yet
- Med Chem 535P - Diagnostic Medicinal Chemistry Hematology Leukocytes (White Blood Cells, WBC)Document9 pagesMed Chem 535P - Diagnostic Medicinal Chemistry Hematology Leukocytes (White Blood Cells, WBC)hadiemNo ratings yet
- Hiv Resistance ClickerDocument28 pagesHiv Resistance ClickerMichael Orekunrin Jr.0% (1)
- Hypersensitivity Reactions: Type III and Type IV HypersensitivityDocument36 pagesHypersensitivity Reactions: Type III and Type IV HypersensitivityDil Navab100% (1)
- Ebook Medical Biochemistry PDF Full Chapter PDFDocument67 pagesEbook Medical Biochemistry PDF Full Chapter PDFfabian.gordon906100% (30)
- Goodpasture's Syndrome, Hashimoto's Thyroiditis, Graves's Disease & Type I DiabetesDocument2 pagesGoodpasture's Syndrome, Hashimoto's Thyroiditis, Graves's Disease & Type I DiabetesZara MohammedNo ratings yet
- Peripherally Induced Brain Tissue-Resident Memory CD8+ T Cells Mediate Protection Against CNS InfectionDocument25 pagesPeripherally Induced Brain Tissue-Resident Memory CD8+ T Cells Mediate Protection Against CNS InfectionFernandoNo ratings yet
- Efector Mechanism of Adaptive Immune ResponseDocument27 pagesEfector Mechanism of Adaptive Immune ResponseHerdian KusumaNo ratings yet
- Immunology Practice Questions With AnswersDocument21 pagesImmunology Practice Questions With AnswersBigBoosting100% (2)
- Acquired ImmunityDocument16 pagesAcquired ImmunityMohammed Hashem100% (1)
- ImmunologyDocument83 pagesImmunologyKühne ZoeNo ratings yet
- Block 1Document108 pagesBlock 1Varshini SrivatsaNo ratings yet
- Specific Immune System: T - LymphocytesDocument18 pagesSpecific Immune System: T - Lymphocytesarun231187No ratings yet
- T CellDocument19 pagesT CellSachin AnandNo ratings yet