Download as pdf or txt
You might also like
- Mô hình kinh doanh thức uống trà Kombucha - Môn quản lý dịch vụ ẩm thựcDocument76 pagesMô hình kinh doanh thức uống trà Kombucha - Môn quản lý dịch vụ ẩm thựcNam NguyenHoangNo ratings yet
- Chuong IIDocument162 pagesChuong IIChâu LêNo ratings yet
- Lec 5Document5 pagesLec 5Hong AnhNo ratings yet
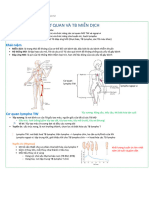
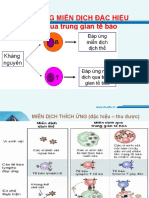
- Kháng Nguyên Và MHCDocument33 pagesKháng Nguyên Và MHCVy HảiNo ratings yet
- 2. TẾ BÀO MIỄN DỊCH VÀ CƠ QUAN LYMPHODocument42 pages2. TẾ BÀO MIỄN DỊCH VÀ CƠ QUAN LYMPHOHoàng DũngNo ratings yet
- De Cuong On Tap Cuoi Ki Mon Mien Dich CompressDocument32 pagesDe Cuong On Tap Cuoi Ki Mon Mien Dich CompressVăn Đường LêNo ratings yet
- Xử lý và trình diện KN-TUYẾN ỨCDocument17 pagesXử lý và trình diện KN-TUYẾN ỨCMinh QuânNo ratings yet
- Miễn dịchDocument4 pagesMiễn dịchHiền SV. Nguyễn Thị ThuNo ratings yet
- Trình Diện Kháng NguyênDocument41 pagesTrình Diện Kháng NguyênMạn Châu Sa HoaNo ratings yet
- Kháng NguyênDocument5 pagesKháng NguyênQuang QuangNo ratings yet
- Chuong 6Document31 pagesChuong 6Thiên Kim LêNo ratings yet
- Bai6 - Te Bao LYMPHO B Va Dap Ung Mien Dich TheDocument12 pagesBai6 - Te Bao LYMPHO B Va Dap Ung Mien Dich Thethamdochucnangtang3No ratings yet
- FILETONGHOPMDDocument30 pagesFILETONGHOPMDngannld12ieltsbmtNo ratings yet
- Lec 3Document15 pagesLec 3Hong AnhNo ratings yet
- LEC 3,4 Huyết họcDocument13 pagesLEC 3,4 Huyết họchocymaivuiNo ratings yet
- Xử Lý Và Trình Diện KN-TUYẾN ỨCDocument17 pagesXử Lý Và Trình Diện KN-TUYẾN ỨCTrung IndusNo ratings yet
- Major Histocompability ComplexDocument63 pagesMajor Histocompability ComplexThi LâmNo ratings yet
- Sinh Bệnh Học Và Đáp Ứng Ký ChủDocument12 pagesSinh Bệnh Học Và Đáp Ứng Ký ChủHiền SV. Nguyễn Thị ThuNo ratings yet
- Đề Cương Miễn DịchDocument11 pagesĐề Cương Miễn Dịch19100071 Nguyễn Minh PhươngNo ratings yet
- MIỄN DỊCHDocument9 pagesMIỄN DỊCHDũng HoàngNo ratings yet
- CÂU HỎI ÔN SINH LÝ BỆNHDocument27 pagesCÂU HỎI ÔN SINH LÝ BỆNHcather12345123No ratings yet
- L.T (SV)Document10 pagesL.T (SV)Quỳnh PhạmNo ratings yet
- 7 Đáp Ứng Miễn Dịch TbDocument28 pages7 Đáp Ứng Miễn Dịch TbQuan LuuNo ratings yet
- Quá Trình Trư NG Thành Và Phân Lo I Lympho TDocument3 pagesQuá Trình Trư NG Thành Và Phân Lo I Lympho TLê Quang NhựtNo ratings yet
- Phần lympho TDocument11 pagesPhần lympho Ttinh906No ratings yet
- chuong 3-Môn miễn dịch học ứng dụngDocument76 pageschuong 3-Môn miễn dịch học ứng dụngNam NguyenHoangNo ratings yet
- DauanbiethoaDocument28 pagesDauanbiethoaPhùngg Văn QuanggNo ratings yet
- Thái Hương Tóm TắtDocument8 pagesThái Hương Tóm Tắtthamdochucnangtang3No ratings yet
- Mien Dich Dich TheDocument23 pagesMien Dich Dich TheWin LâmNo ratings yet
- MD SLB 2020 2021 Copy 1Document13 pagesMD SLB 2020 2021 Copy 1Lee Eun YeonNo ratings yet
- KN Va MHC - Hua.2020Document30 pagesKN Va MHC - Hua.2020Nguyễn QuânNo ratings yet
- Miễn dịchDocument2 pagesMiễn dịchnhoangminh961No ratings yet
- Hệ miễn dịchDocument27 pagesHệ miễn dịchHuỳnh Tiến KhoaNo ratings yet
- MIỄN DỊCH THẦY TIẾNDocument50 pagesMIỄN DỊCH THẦY TIẾNdonguyenmaitrinh2005No ratings yet
- MD Thich NghiDocument37 pagesMD Thich NghiMạn Châu Sa HoaNo ratings yet
- 2. Tổ chức và tế bào của hệ MDDocument14 pages2. Tổ chức và tế bào của hệ MDĐậu MầmNo ratings yet
- Bài 3 Sinh lý đại cương miễn dịchDocument53 pagesBài 3 Sinh lý đại cương miễn dịchtranla.mrNo ratings yet
- P. Thúc ĐƯMD (SV)Document14 pagesP. Thúc ĐƯMD (SV)Quỳnh PhạmNo ratings yet
- Dòng Lympho TDocument28 pagesDòng Lympho TThành Nguyễn VănNo ratings yet
- B. 3 - KHÁNG NGUYÊN - KHÁNG THỂDocument25 pagesB. 3 - KHÁNG NGUYÊN - KHÁNG THỂshinichi kudoNo ratings yet
- Miễn DịchDocument20 pagesMiễn Dịchnguyenngoc2000nekNo ratings yet
- B11. Khái niệm đáp ứng MDDocument46 pagesB11. Khái niệm đáp ứng MDNhung NguyễnNo ratings yet
- Dong Hoc Dap Ung Mien DichDocument24 pagesDong Hoc Dap Ung Mien DichHuy GiaNo ratings yet
- HỆ BẠCH HUYẾT - HỆ MIỄN DỊCHDocument29 pagesHỆ BẠCH HUYẾT - HỆ MIỄN DỊCHSangg Đức100% (2)
- Sli BCDocument3 pagesSli BChoangmy2314No ratings yet
- Chức năng của Kháng ThểDocument3 pagesChức năng của Kháng ThểĐăngNo ratings yet
- hand out u lympho vung hoc mieng- TS Phan Đặng Anh ThưDocument34 pageshand out u lympho vung hoc mieng- TS Phan Đặng Anh ThưThế anh PhạmNo ratings yet
- Mien DichDocument4 pagesMien DichĐỗ Nguyệt HàNo ratings yet
- L.B (SV)Document10 pagesL.B (SV)Quỳnh PhạmNo ratings yet
- Chuong 1+2-Đã NénDocument37 pagesChuong 1+2-Đã NénPhan NguyễnNo ratings yet
- +Provirut nằm dưới sự kiểm soát của tế bào chủ và phiên mã nhờ ARN polimeraza II của tế bàoDocument2 pages+Provirut nằm dưới sự kiểm soát của tế bào chủ và phiên mã nhờ ARN polimeraza II của tế bàoduykhanhnguyen3528No ratings yet
- MD SLB 21 22 Duoc Kì PHDocument8 pagesMD SLB 21 22 Duoc Kì PHHoài PhươngNo ratings yet
- Chuong IDocument74 pagesChuong Inguyentnhu9522No ratings yet
- CQ Tao Huyet Va Mien DichDocument47 pagesCQ Tao Huyet Va Mien DichLý Nguyễn Chiến ThắngNo ratings yet
- Chương 5-Kháng Thể Dịch Thể Đặc HiệuDocument73 pagesChương 5-Kháng Thể Dịch Thể Đặc HiệuthuyvyhnkNo ratings yet
- VI Sinh-NttuDocument3 pagesVI Sinh-NttuTú AnhNo ratings yet
- Dung nạp miễn dịch và tự miễnDocument57 pagesDung nạp miễn dịch và tự miễnPhương AnhNo ratings yet
- Cơ Quan Miễn DịchDocument17 pagesCơ Quan Miễn DịchPhạm OanhNo ratings yet
- 5. BẠCH CẦU ĐƠN NHÂN 2021Document62 pages5. BẠCH CẦU ĐƠN NHÂN 2021Nguyễn Sơn TùngNo ratings yet
- Tổng Quan Các Chất Độc Acrylamide, PAHs, Furan Trong Chiên Nướng Từ Các Sản Phẩm Tinh Bột- IUHDocument60 pagesTổng Quan Các Chất Độc Acrylamide, PAHs, Furan Trong Chiên Nướng Từ Các Sản Phẩm Tinh Bột- IUHNam NguyenHoangNo ratings yet
- Ảnh hưởng của trạng thái lỏng, rắn của thực phẩm đối với lượng thức ăn do con người dung nạp- IUHDocument40 pagesẢnh hưởng của trạng thái lỏng, rắn của thực phẩm đối với lượng thức ăn do con người dung nạp- IUHNam NguyenHoangNo ratings yet
- Đề Thi Cuối Kì Môn Dinh Dưỡng Lâm Sàng- IUHDocument1 pageĐề Thi Cuối Kì Môn Dinh Dưỡng Lâm Sàng- IUHNam NguyenHoangNo ratings yet
- Tổng quan về mối liên hệ giữa dinh dưỡng và di truyền học biểu sinh- IUHDocument41 pagesTổng quan về mối liên hệ giữa dinh dưỡng và di truyền học biểu sinh- IUHNam NguyenHoangNo ratings yet
- Vai trò của vận động trong quản lý thừa cân béo phì- Học thuật với chuyên gia lần 2Document59 pagesVai trò của vận động trong quản lý thừa cân béo phì- Học thuật với chuyên gia lần 2Nam NguyenHoangNo ratings yet
- Xây dựng hệ thống Haccp- Snack khoai tây vị phô mai KracksDocument15 pagesXây dựng hệ thống Haccp- Snack khoai tây vị phô mai KracksNam NguyenHoangNo ratings yet
- Tổng hợp kiến thức về di truyền học- https://suckhoeditruyen.vnDocument67 pagesTổng hợp kiến thức về di truyền học- https://suckhoeditruyen.vnNam NguyenHoang100% (1)
- XƯƠNG - THẤP KHỚP N9Document38 pagesXƯƠNG - THẤP KHỚP N9Nam NguyenHoangNo ratings yet
- Xác định độ tinh khiết tinh dầu- Môn hóa học hợp chất tự nhiênDocument9 pagesXác định độ tinh khiết tinh dầu- Môn hóa học hợp chất tự nhiênNam NguyenHoangNo ratings yet
- Xây dựng hệ thống Haccp- chuối sấy dẻoDocument16 pagesXây dựng hệ thống Haccp- chuối sấy dẻoNam NguyenHoangNo ratings yet
- Chương 5 - Môn thiết kế và phát triển sản phẩm - IUHDocument11 pagesChương 5 - Môn thiết kế và phát triển sản phẩm - IUHNam NguyenHoangNo ratings yet
- 2021 Mau Khoa Luan Tot NghiepDocument19 pages2021 Mau Khoa Luan Tot NghiepGiang Phạm HuỳnhNo ratings yet
- Nước ép trái cây kết hợp với rau, củ- Môn thiết kế và phát triển sản phẩm- IUHDocument41 pagesNước ép trái cây kết hợp với rau, củ- Môn thiết kế và phát triển sản phẩm- IUHNam NguyenHoangNo ratings yet
- Bài báo cáo thực hành môn công nghệ chế biến nông sản- IUHDocument35 pagesBài báo cáo thực hành môn công nghệ chế biến nông sản- IUHNam NguyenHoangNo ratings yet
- Chương 1 - Môn thiết kế và phát triển sản phẩm - IUHDocument22 pagesChương 1 - Môn thiết kế và phát triển sản phẩm - IUHNam NguyenHoangNo ratings yet
- Chương 2- Môn thiết kế và phát triển sản phẩm - IUHDocument16 pagesChương 2- Môn thiết kế và phát triển sản phẩm - IUHNam NguyenHoangNo ratings yet
- Liệu pháp dinh dưỡng cho đối tượng bị rối loạn gan mật - Dinh dưỡng lâm sàngDocument48 pagesLiệu pháp dinh dưỡng cho đối tượng bị rối loạn gan mật - Dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet
- Chương 3- Môn thiết kế và phát triển sản phẩm - IUHDocument27 pagesChương 3- Môn thiết kế và phát triển sản phẩm - IUHNam NguyenHoangNo ratings yet
- Chương 4 - Môn thiết kế và phát triển sản phẩm - IUHDocument16 pagesChương 4 - Môn thiết kế và phát triển sản phẩm - IUHNam NguyenHoangNo ratings yet
- PPT thuyết trình môn chế biến thực dưỡng - IUHDocument29 pagesPPT thuyết trình môn chế biến thực dưỡng - IUHNam NguyenHoangNo ratings yet
- Béo Phì Ngư I L N - Môn Dinh Dư NG Lâm SàngDocument25 pagesBéo Phì Ngư I L N - Môn Dinh Dư NG Lâm SàngNam NguyenHoangNo ratings yet
- Đại cương môn tin học - IUHDocument863 pagesĐại cương môn tin học - IUHNam NguyenHoangNo ratings yet
- Bệnh thận và dinh dưỡng điều trị - Môn dinh dưỡng lâm sàngDocument27 pagesBệnh thận và dinh dưỡng điều trị - Môn dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet
- Thiết kế mô hình kinh doanh các loại bánh quy yến và trà nhằm hạn chế bệnh tiểu đường- Môn quản lý dịch vụ ẩm thựcDocument59 pagesThiết kế mô hình kinh doanh các loại bánh quy yến và trà nhằm hạn chế bệnh tiểu đường- Môn quản lý dịch vụ ẩm thựcNam NguyenHoangNo ratings yet
- Bỏng và phẫu - Phòng ngừa và điều trị ung thư- Môn dinh dưỡng lâm sàngDocument38 pagesBỏng và phẫu - Phòng ngừa và điều trị ung thư- Môn dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet
- Bệnh tim mạch- Môn dinh dưỡng lâm sàngDocument43 pagesBệnh tim mạch- Môn dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet
- Quản lý cân nặng cho bệnh nhân thừa thừa - béo phì, suy dinh dưỡng- Môn dinh dưỡng lâm sàngDocument35 pagesQuản lý cân nặng cho bệnh nhân thừa thừa - béo phì, suy dinh dưỡng- Môn dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet
- Bệnh rối loạn tiêu hóa - word - Môn dinh dưỡng lâm sàngDocument33 pagesBệnh rối loạn tiêu hóa - word - Môn dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet
- Bệnh rối loạn tiêu hóa - ppt- Môn dinh dưỡng lâm sàngDocument42 pagesBệnh rối loạn tiêu hóa - ppt- Môn dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet