
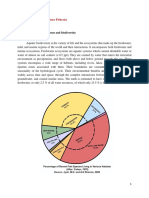
The Freshwater Habitats Fishes and Fisheries of TH
The Freshwater Habitats Fishes and Fisheries of TH
You might also like
- The Biologist 2018 16 1 139-158 Franco-Lopez EtalDocument20 pagesThe Biologist 2018 16 1 139-158 Franco-Lopez EtalIlver AlabatNo ratings yet
- Diversity and Distribution of Freshwater Fish PDFDocument13 pagesDiversity and Distribution of Freshwater Fish PDFBob UrbandubNo ratings yet
- Bio323review Paper 2Document10 pagesBio323review Paper 2kashi1994No ratings yet
- Session 3 Review River Fisheries: Ecological Basis For Management and ConservationDocument40 pagesSession 3 Review River Fisheries: Ecological Basis For Management and ConservationLove RebiraNo ratings yet
- Ecosystem of Chilka LakeDocument10 pagesEcosystem of Chilka LakeSachin Kumar BassiNo ratings yet
- 2011 Rodriguez-Olarte Et Al Fish Status Abrae VenezuelaDocument13 pages2011 Rodriguez-Olarte Et Al Fish Status Abrae VenezuelaDouglas Rodriguez OlarteNo ratings yet
- Carvalho, Zuanon, Sazima EOLSSDocument32 pagesCarvalho, Zuanon, Sazima EOLSSranitapyNo ratings yet
- Amphibians and Reptiles of Luzon Island, V: The Herpetofauna of Angat Dam Watershed, Bulacan Province, Luzon Island, PhilippinesDocument23 pagesAmphibians and Reptiles of Luzon Island, V: The Herpetofauna of Angat Dam Watershed, Bulacan Province, Luzon Island, PhilippinesLakber MandNo ratings yet
- Harmful AlgaDocument20 pagesHarmful AlgaAsa Étudier La-DienNo ratings yet
- Biodiversity Index of Corals in Bitaug StA. Cruz Davao Del SurDocument74 pagesBiodiversity Index of Corals in Bitaug StA. Cruz Davao Del SurMicky BanderaNo ratings yet
- Art 11Document13 pagesArt 11Jeancarlos QuispeNo ratings yet
- Chilvers Et AlDocument13 pagesChilvers Et Alapi-3828346No ratings yet
- Tarazona Et Al. 2003 Overview and Challenges of Marine BiodiversityDocument26 pagesTarazona Et Al. 2003 Overview and Challenges of Marine BiodiversitySonia ValleNo ratings yet
- Whit Field, 1999) Ichthyofaunal Assemblages in Estuaries - A South African Case StudyDocument36 pagesWhit Field, 1999) Ichthyofaunal Assemblages in Estuaries - A South African Case StudyAndré S. FilguerasNo ratings yet
- Cohen & Carlton 1995 San Francisco Bay Invasion ReportDocument292 pagesCohen & Carlton 1995 San Francisco Bay Invasion Reportacohen321No ratings yet
- Freshwater Fish in SabahDocument9 pagesFreshwater Fish in SabahSteve NgNo ratings yet
- Fish and Fisheries of The Eastern Coast of MexicoDocument20 pagesFish and Fisheries of The Eastern Coast of MexicoKel'Thu Freking'ZadNo ratings yet
- SCRIBD2Document14 pagesSCRIBD2cesar8319No ratings yet
- Aquatic Ecosystem Health & ManagementDocument22 pagesAquatic Ecosystem Health & ManagementAlessandra MendonçaNo ratings yet
- Heterogeneidad Ambiental y Su Relación Con La Diversidad y AbundanciaDocument20 pagesHeterogeneidad Ambiental y Su Relación Con La Diversidad y AbundanciaJuan PerezNo ratings yet
- BFSC 205 - Inland FisheriesDocument177 pagesBFSC 205 - Inland FisheriesSruthiNo ratings yet
- Marine Ecosystems Are Among The Largest of Earth'sDocument5 pagesMarine Ecosystems Are Among The Largest of Earth'skevinaveriaNo ratings yet
- Species Composition of Macroinvertebrates in Sto. Tomas Cove, La Union, PhilippinesDocument10 pagesSpecies Composition of Macroinvertebrates in Sto. Tomas Cove, La Union, PhilippinesJfkfkfjfNo ratings yet
- 1.1 Background of The StudyDocument27 pages1.1 Background of The StudyLeo JahackNo ratings yet
- Dams and The Fish Fauna of The Neotropical Region: Impacts and Management Related To Diversity and FisheriesDocument14 pagesDams and The Fish Fauna of The Neotropical Region: Impacts and Management Related To Diversity and FisheriesmarceloNo ratings yet
- Fish MigrationDocument23 pagesFish MigrationSukanya DeviNo ratings yet
- Jcoastres D 16 00155.1Document13 pagesJcoastres D 16 00155.1valentina vargasNo ratings yet
- Abundance and Diversity of Fishes in BayDocument17 pagesAbundance and Diversity of Fishes in BayRedgie PotenciandoNo ratings yet
- Gunkel, 2000Document8 pagesGunkel, 2000Fernanda Egas GaeteNo ratings yet
- Introduction To The Caura River Basin, Bolívar State, VenezuelaDocument7 pagesIntroduction To The Caura River Basin, Bolívar State, VenezuelaEnrique MartinezNo ratings yet
- Aquatic Biodiversity of Lake Mainit, Southern PhilippinesDocument12 pagesAquatic Biodiversity of Lake Mainit, Southern PhilippinesRICA MAE LUMICTINNo ratings yet
- Asociaciones PecesDocument13 pagesAsociaciones PecesjorgelopezrochaNo ratings yet
- Doradoetal 2021 TaxDivandEcoAttribofCopesfromCol NeotBiodDocument14 pagesDoradoetal 2021 TaxDivandEcoAttribofCopesfromCol NeotBiodTvbox JiménezNo ratings yet
- Overview of FishDocument20 pagesOverview of FishCHAN KIMSYNo ratings yet
- 46_SanchezVelasco_etal_2022_JMarSysDocument10 pages46_SanchezVelasco_etal_2022_JMarSystherion66No ratings yet
- Estructura Trófica de Los Peces en Arroyos Del Corral de San Luis, Cuenca Del Bajo Magdalena, Caribe, ColombiaDocument19 pagesEstructura Trófica de Los Peces en Arroyos Del Corral de San Luis, Cuenca Del Bajo Magdalena, Caribe, ColombiaBaltazar Calderon RoncalloNo ratings yet
- Lecture For Fisheries 2014 Harmonised For StudentDocument134 pagesLecture For Fisheries 2014 Harmonised For StudentnataniumcscbeNo ratings yet
- Mascareñas-Osorio Et Al. - 2011 - Checklist of Conspicuous Reef Fishes of The Bahía PDFDocument9 pagesMascareñas-Osorio Et Al. - 2011 - Checklist of Conspicuous Reef Fishes of The Bahía PDFDaniel OviedoNo ratings yet
- Abundancia, Tamaño y Estructura Poblacional Del Tiburón Punta Blanca de Arrecife, Triaenodon Obesus (Carcharhiniformes: Carcharhinidae), en Bahía Chatham, Parque Nacional Isla Del Coco, Costa RicaDocument8 pagesAbundancia, Tamaño y Estructura Poblacional Del Tiburón Punta Blanca de Arrecife, Triaenodon Obesus (Carcharhiniformes: Carcharhinidae), en Bahía Chatham, Parque Nacional Isla Del Coco, Costa RicaDayanna MeraNo ratings yet
- Ecosistemas Coralinos Del Área de Conservación Osa, Costa Rica - Esttructura y Necesidades de ConservaciónDocument42 pagesEcosistemas Coralinos Del Área de Conservación Osa, Costa Rica - Esttructura y Necesidades de ConservaciónJuanNo ratings yet
- Physics and Chemistry of The Earth: I. Naigaga, H. Kaiser, W.J. Muller, L. Ojok, D. Mbabazi, G. Magezi, E. MuhumuzaDocument11 pagesPhysics and Chemistry of The Earth: I. Naigaga, H. Kaiser, W.J. Muller, L. Ojok, D. Mbabazi, G. Magezi, E. MuhumuzaAngela JuliethNo ratings yet
- Williner Et Al 2014Document9 pagesWilliner Et Al 2014veronica willinerNo ratings yet
- Structural Changes in Freshwater Mussel Bivalviaunionidae Assemblages Downstream of Lake Somerville TexasDocument9 pagesStructural Changes in Freshwater Mussel Bivalviaunionidae Assemblages Downstream of Lake Somerville TexasfrancislouisemaduramenteNo ratings yet
- Potential Effects of Runoff, Fluvial Sediment, and Nutrient Discharges On The Coral Reefs of Puerto Rico 36285527Document21 pagesPotential Effects of Runoff, Fluvial Sediment, and Nutrient Discharges On The Coral Reefs of Puerto Rico 36285527Anthony McIvorNo ratings yet
- Fish Zonations and Guilds As The Basis For Assessment of Ecological Integrity of Large RiversDocument22 pagesFish Zonations and Guilds As The Basis For Assessment of Ecological Integrity of Large RiversPaula Rendón CardonaNo ratings yet
- Influence of Water Quality On The Biodiversity ofDocument7 pagesInfluence of Water Quality On The Biodiversity ofFor NewslettersNo ratings yet
- Stdprod 0858361Document88 pagesStdprod 0858361mcoada82No ratings yet
- Alcocer - 1998 - Saline Lake Ecosystems of MexicoDocument26 pagesAlcocer - 1998 - Saline Lake Ecosystems of Mexicogrammerly20 2017No ratings yet
- AkomaImoobepaperDocument9 pagesAkomaImoobepaperdegseraNo ratings yet
- Araya Et Al 2014 - Migracion Parcila de ChinookDocument9 pagesAraya Et Al 2014 - Migracion Parcila de ChinookRodrigo Vera S.No ratings yet
- Fishers' Local Ecological Knowledge Indicate Migration Patterns of Tropical Freshwater Fish in An Amazonian RiverDocument19 pagesFishers' Local Ecological Knowledge Indicate Migration Patterns of Tropical Freshwater Fish in An Amazonian RiversestefanieNo ratings yet
- Willamette Salmon HabitatDocument21 pagesWillamette Salmon HabitatKGW News100% (1)
- Estructuratrficadelospecesenarroyosdel Corralde San Luis 2016Document21 pagesEstructuratrficadelospecesenarroyosdel Corralde San Luis 2016Baltazar Calderon RoncalloNo ratings yet
- Articulo PDFDocument9 pagesArticulo PDFDaniel Miguel Manchego RamosNo ratings yet
- E Ffects of Fishing On The Structure and Functioning of Estuarine and Nearshore EcosystemsDocument13 pagesE Ffects of Fishing On The Structure and Functioning of Estuarine and Nearshore EcosystemsnajwamohamadNo ratings yet
- TUGAS 08 Pengetahuan Fisik Laut'Document6 pagesTUGAS 08 Pengetahuan Fisik Laut'Rizaldi CaesarNo ratings yet
- Diversity: Macroinvertebrate Taxonomic Richness in Minimally Disturbed Streams On The Southeastern USA Coastal PlainDocument17 pagesDiversity: Macroinvertebrate Taxonomic Richness in Minimally Disturbed Streams On The Southeastern USA Coastal PlainViviana BravoNo ratings yet
- Marine Inverteb-WPS OfficeDocument22 pagesMarine Inverteb-WPS OfficembojefadhilNo ratings yet
- Fortes Et Al 2013Document12 pagesFortes Et Al 2013monteironetocassNo ratings yet
- Floodplain Wetland Biota in the Murray-Darling Basin: Water and Habitat RequirementsFrom EverandFloodplain Wetland Biota in the Murray-Darling Basin: Water and Habitat RequirementsKerrylee RogersNo ratings yet
- Shallow Groundwater Quality of Lahore City Along The River Ravi by Afaf AyeshaDocument9 pagesShallow Groundwater Quality of Lahore City Along The River Ravi by Afaf AyeshaShumaila MajeedNo ratings yet
- Assignment 1 Disaster& Risk Management Submitted By: Ezza Ahmad Butt Submitted To: Ma'Am Shumaila Submission Date: 24 SPET 2021 Semester 7Document5 pagesAssignment 1 Disaster& Risk Management Submitted By: Ezza Ahmad Butt Submitted To: Ma'Am Shumaila Submission Date: 24 SPET 2021 Semester 7Shumaila MajeedNo ratings yet
- 1 Groundwater Levels, Temperature and QualityDocument14 pages1 Groundwater Levels, Temperature and QualityShumaila MajeedNo ratings yet
- Droughts: Types of DroughtDocument3 pagesDroughts: Types of DroughtShumaila MajeedNo ratings yet
- Metropolitian Drain of Lahore CityDocument15 pagesMetropolitian Drain of Lahore CityShumaila MajeedNo ratings yet
- DLL January 9, 2024Document4 pagesDLL January 9, 2024Jezreel RabaNo ratings yet
- Narrative PHT3Document3 pagesNarrative PHT3Jericho Basmayor SajulanNo ratings yet
- Indiv Scores April24th & 25th 2010Document4 pagesIndiv Scores April24th & 25th 2010ABESC150No ratings yet
- Survive Mola Mola - OrvDocument35 pagesSurvive Mola Mola - OrvPhương Lương MaiNo ratings yet
- Aqua History PhilDocument27 pagesAqua History Philvirgil mae taghapNo ratings yet
- 4th Science Notes The HopeDocument67 pages4th Science Notes The Hopemasterm9800No ratings yet
- Calantas, Jana Karla C. BS Fisheries III: Land-Based Hatchery and Juvenile Production FarmDocument3 pagesCalantas, Jana Karla C. BS Fisheries III: Land-Based Hatchery and Juvenile Production FarmJana Karla CalantasNo ratings yet
- Catfish Farming Information Guide - AgrifarmingDocument5 pagesCatfish Farming Information Guide - AgrifarmingamolkajaleNo ratings yet
- Fisheries BiologyDocument29 pagesFisheries Biologybikashh100% (3)
- Theodoropoulou - Tuna Fishing in The Aegean-2013Document1 pageTheodoropoulou - Tuna Fishing in The Aegean-2013TeSoTrasNo ratings yet
- Chilled and Frozen Raw Fish PDFDocument26 pagesChilled and Frozen Raw Fish PDFMourad AttouchiNo ratings yet
- AquariumDocument65 pagesAquariumDaisy100% (4)
- PBS Baits E - Book Version 1Document48 pagesPBS Baits E - Book Version 1Dirk Henn0% (1)
- African Catfish (Clarias Gariepinus) : Introduction To African Catfish Hatchery Set Up and ManagementDocument14 pagesAfrican Catfish (Clarias Gariepinus) : Introduction To African Catfish Hatchery Set Up and ManagementHaymanot yenesewNo ratings yet
- International Standard Statistical Classification of Fishery Vessels by Vessel Types (ISSCFV, Rev.1, 2019)Document3 pagesInternational Standard Statistical Classification of Fishery Vessels by Vessel Types (ISSCFV, Rev.1, 2019)okta bedjoNo ratings yet
- What Is The Scope of Aquaculture?Document2 pagesWhat Is The Scope of Aquaculture?Louie YamonNo ratings yet
- Proximate Composition of Some Commercial Fish Feeds Sold in NigeriaDocument5 pagesProximate Composition of Some Commercial Fish Feeds Sold in NigeriaDR Ebthal GaberNo ratings yet
- Workshop On Assessment and Management of The Offshore Resources of South and Southeast AsiaDocument44 pagesWorkshop On Assessment and Management of The Offshore Resources of South and Southeast AsiadutvaNo ratings yet
- Optimalisasi Pemanfaatan Dan Pengelolaan Pangkalan Pendaratan Ikan (Ppi) Klaligi Kota Sorong Propinsi Papua BaratDocument150 pagesOptimalisasi Pemanfaatan Dan Pengelolaan Pangkalan Pendaratan Ikan (Ppi) Klaligi Kota Sorong Propinsi Papua BaratMuhammad IqbalNo ratings yet
- Use of Azolla Pinnata (RRL)Document9 pagesUse of Azolla Pinnata (RRL)Jennie Marlow AddisonNo ratings yet
- World Kids Colouring Day Magazine 2016Document12 pagesWorld Kids Colouring Day Magazine 2016Carmen DraghiaNo ratings yet
- 6120838371344Document6 pages6120838371344DeeNo ratings yet
- Fish Processing: Santiago CampusDocument24 pagesFish Processing: Santiago CampusRomnick Elgamo GalantoNo ratings yet
- Endemic Fish of The Birds Head - G. AllenDocument5 pagesEndemic Fish of The Birds Head - G. AllenRudy PriatnaNo ratings yet
- Learning Activity Sheet No.1 in Science 2: Key ConceptDocument25 pagesLearning Activity Sheet No.1 in Science 2: Key ConceptMary Grace BaldozaNo ratings yet
- Fish and Shellfish IdentificationDocument107 pagesFish and Shellfish IdentificationthewurzelNo ratings yet
- +1 Bio Zoo em Vol 1Document30 pages+1 Bio Zoo em Vol 1gowthamgokul206No ratings yet
- Elementary Group Pop-Quiz 115 ReadingDocument3 pagesElementary Group Pop-Quiz 115 ReadingChristophe Kodjo CharlecNo ratings yet
- Fish Species NotebookDocument20 pagesFish Species NotebookLead HeadNo ratings yet
- 2nd Quarter Assessment in Cookery 10Document3 pages2nd Quarter Assessment in Cookery 10Irish Borga100% (1)





























































































Download as pdf or txt
You might also like
- The Biologist 2018 16 1 139-158 Franco-Lopez EtalDocument20 pagesThe Biologist 2018 16 1 139-158 Franco-Lopez EtalIlver AlabatNo ratings yet
- Diversity and Distribution of Freshwater Fish PDFDocument13 pagesDiversity and Distribution of Freshwater Fish PDFBob UrbandubNo ratings yet
- Bio323review Paper 2Document10 pagesBio323review Paper 2kashi1994No ratings yet
- Session 3 Review River Fisheries: Ecological Basis For Management and ConservationDocument40 pagesSession 3 Review River Fisheries: Ecological Basis For Management and ConservationLove RebiraNo ratings yet
- Ecosystem of Chilka LakeDocument10 pagesEcosystem of Chilka LakeSachin Kumar BassiNo ratings yet
- 2011 Rodriguez-Olarte Et Al Fish Status Abrae VenezuelaDocument13 pages2011 Rodriguez-Olarte Et Al Fish Status Abrae VenezuelaDouglas Rodriguez OlarteNo ratings yet
- Carvalho, Zuanon, Sazima EOLSSDocument32 pagesCarvalho, Zuanon, Sazima EOLSSranitapyNo ratings yet
- Amphibians and Reptiles of Luzon Island, V: The Herpetofauna of Angat Dam Watershed, Bulacan Province, Luzon Island, PhilippinesDocument23 pagesAmphibians and Reptiles of Luzon Island, V: The Herpetofauna of Angat Dam Watershed, Bulacan Province, Luzon Island, PhilippinesLakber MandNo ratings yet
- Harmful AlgaDocument20 pagesHarmful AlgaAsa Étudier La-DienNo ratings yet
- Biodiversity Index of Corals in Bitaug StA. Cruz Davao Del SurDocument74 pagesBiodiversity Index of Corals in Bitaug StA. Cruz Davao Del SurMicky BanderaNo ratings yet
- Art 11Document13 pagesArt 11Jeancarlos QuispeNo ratings yet
- Chilvers Et AlDocument13 pagesChilvers Et Alapi-3828346No ratings yet
- Tarazona Et Al. 2003 Overview and Challenges of Marine BiodiversityDocument26 pagesTarazona Et Al. 2003 Overview and Challenges of Marine BiodiversitySonia ValleNo ratings yet
- Whit Field, 1999) Ichthyofaunal Assemblages in Estuaries - A South African Case StudyDocument36 pagesWhit Field, 1999) Ichthyofaunal Assemblages in Estuaries - A South African Case StudyAndré S. FilguerasNo ratings yet
- Cohen & Carlton 1995 San Francisco Bay Invasion ReportDocument292 pagesCohen & Carlton 1995 San Francisco Bay Invasion Reportacohen321No ratings yet
- Freshwater Fish in SabahDocument9 pagesFreshwater Fish in SabahSteve NgNo ratings yet
- Fish and Fisheries of The Eastern Coast of MexicoDocument20 pagesFish and Fisheries of The Eastern Coast of MexicoKel'Thu Freking'ZadNo ratings yet
- SCRIBD2Document14 pagesSCRIBD2cesar8319No ratings yet
- Aquatic Ecosystem Health & ManagementDocument22 pagesAquatic Ecosystem Health & ManagementAlessandra MendonçaNo ratings yet
- Heterogeneidad Ambiental y Su Relación Con La Diversidad y AbundanciaDocument20 pagesHeterogeneidad Ambiental y Su Relación Con La Diversidad y AbundanciaJuan PerezNo ratings yet
- BFSC 205 - Inland FisheriesDocument177 pagesBFSC 205 - Inland FisheriesSruthiNo ratings yet
- Marine Ecosystems Are Among The Largest of Earth'sDocument5 pagesMarine Ecosystems Are Among The Largest of Earth'skevinaveriaNo ratings yet
- Species Composition of Macroinvertebrates in Sto. Tomas Cove, La Union, PhilippinesDocument10 pagesSpecies Composition of Macroinvertebrates in Sto. Tomas Cove, La Union, PhilippinesJfkfkfjfNo ratings yet
- 1.1 Background of The StudyDocument27 pages1.1 Background of The StudyLeo JahackNo ratings yet
- Dams and The Fish Fauna of The Neotropical Region: Impacts and Management Related To Diversity and FisheriesDocument14 pagesDams and The Fish Fauna of The Neotropical Region: Impacts and Management Related To Diversity and FisheriesmarceloNo ratings yet
- Fish MigrationDocument23 pagesFish MigrationSukanya DeviNo ratings yet
- Jcoastres D 16 00155.1Document13 pagesJcoastres D 16 00155.1valentina vargasNo ratings yet
- Abundance and Diversity of Fishes in BayDocument17 pagesAbundance and Diversity of Fishes in BayRedgie PotenciandoNo ratings yet
- Gunkel, 2000Document8 pagesGunkel, 2000Fernanda Egas GaeteNo ratings yet
- Introduction To The Caura River Basin, Bolívar State, VenezuelaDocument7 pagesIntroduction To The Caura River Basin, Bolívar State, VenezuelaEnrique MartinezNo ratings yet
- Aquatic Biodiversity of Lake Mainit, Southern PhilippinesDocument12 pagesAquatic Biodiversity of Lake Mainit, Southern PhilippinesRICA MAE LUMICTINNo ratings yet
- Asociaciones PecesDocument13 pagesAsociaciones PecesjorgelopezrochaNo ratings yet
- Doradoetal 2021 TaxDivandEcoAttribofCopesfromCol NeotBiodDocument14 pagesDoradoetal 2021 TaxDivandEcoAttribofCopesfromCol NeotBiodTvbox JiménezNo ratings yet
- Overview of FishDocument20 pagesOverview of FishCHAN KIMSYNo ratings yet
- 46_SanchezVelasco_etal_2022_JMarSysDocument10 pages46_SanchezVelasco_etal_2022_JMarSystherion66No ratings yet
- Estructura Trófica de Los Peces en Arroyos Del Corral de San Luis, Cuenca Del Bajo Magdalena, Caribe, ColombiaDocument19 pagesEstructura Trófica de Los Peces en Arroyos Del Corral de San Luis, Cuenca Del Bajo Magdalena, Caribe, ColombiaBaltazar Calderon RoncalloNo ratings yet
- Lecture For Fisheries 2014 Harmonised For StudentDocument134 pagesLecture For Fisheries 2014 Harmonised For StudentnataniumcscbeNo ratings yet
- Mascareñas-Osorio Et Al. - 2011 - Checklist of Conspicuous Reef Fishes of The Bahía PDFDocument9 pagesMascareñas-Osorio Et Al. - 2011 - Checklist of Conspicuous Reef Fishes of The Bahía PDFDaniel OviedoNo ratings yet
- Abundancia, Tamaño y Estructura Poblacional Del Tiburón Punta Blanca de Arrecife, Triaenodon Obesus (Carcharhiniformes: Carcharhinidae), en Bahía Chatham, Parque Nacional Isla Del Coco, Costa RicaDocument8 pagesAbundancia, Tamaño y Estructura Poblacional Del Tiburón Punta Blanca de Arrecife, Triaenodon Obesus (Carcharhiniformes: Carcharhinidae), en Bahía Chatham, Parque Nacional Isla Del Coco, Costa RicaDayanna MeraNo ratings yet
- Ecosistemas Coralinos Del Área de Conservación Osa, Costa Rica - Esttructura y Necesidades de ConservaciónDocument42 pagesEcosistemas Coralinos Del Área de Conservación Osa, Costa Rica - Esttructura y Necesidades de ConservaciónJuanNo ratings yet
- Physics and Chemistry of The Earth: I. Naigaga, H. Kaiser, W.J. Muller, L. Ojok, D. Mbabazi, G. Magezi, E. MuhumuzaDocument11 pagesPhysics and Chemistry of The Earth: I. Naigaga, H. Kaiser, W.J. Muller, L. Ojok, D. Mbabazi, G. Magezi, E. MuhumuzaAngela JuliethNo ratings yet
- Williner Et Al 2014Document9 pagesWilliner Et Al 2014veronica willinerNo ratings yet
- Structural Changes in Freshwater Mussel Bivalviaunionidae Assemblages Downstream of Lake Somerville TexasDocument9 pagesStructural Changes in Freshwater Mussel Bivalviaunionidae Assemblages Downstream of Lake Somerville TexasfrancislouisemaduramenteNo ratings yet
- Potential Effects of Runoff, Fluvial Sediment, and Nutrient Discharges On The Coral Reefs of Puerto Rico 36285527Document21 pagesPotential Effects of Runoff, Fluvial Sediment, and Nutrient Discharges On The Coral Reefs of Puerto Rico 36285527Anthony McIvorNo ratings yet
- Fish Zonations and Guilds As The Basis For Assessment of Ecological Integrity of Large RiversDocument22 pagesFish Zonations and Guilds As The Basis For Assessment of Ecological Integrity of Large RiversPaula Rendón CardonaNo ratings yet
- Influence of Water Quality On The Biodiversity ofDocument7 pagesInfluence of Water Quality On The Biodiversity ofFor NewslettersNo ratings yet
- Stdprod 0858361Document88 pagesStdprod 0858361mcoada82No ratings yet
- Alcocer - 1998 - Saline Lake Ecosystems of MexicoDocument26 pagesAlcocer - 1998 - Saline Lake Ecosystems of Mexicogrammerly20 2017No ratings yet
- AkomaImoobepaperDocument9 pagesAkomaImoobepaperdegseraNo ratings yet
- Araya Et Al 2014 - Migracion Parcila de ChinookDocument9 pagesAraya Et Al 2014 - Migracion Parcila de ChinookRodrigo Vera S.No ratings yet
- Fishers' Local Ecological Knowledge Indicate Migration Patterns of Tropical Freshwater Fish in An Amazonian RiverDocument19 pagesFishers' Local Ecological Knowledge Indicate Migration Patterns of Tropical Freshwater Fish in An Amazonian RiversestefanieNo ratings yet
- Willamette Salmon HabitatDocument21 pagesWillamette Salmon HabitatKGW News100% (1)
- Estructuratrficadelospecesenarroyosdel Corralde San Luis 2016Document21 pagesEstructuratrficadelospecesenarroyosdel Corralde San Luis 2016Baltazar Calderon RoncalloNo ratings yet
- Articulo PDFDocument9 pagesArticulo PDFDaniel Miguel Manchego RamosNo ratings yet
- E Ffects of Fishing On The Structure and Functioning of Estuarine and Nearshore EcosystemsDocument13 pagesE Ffects of Fishing On The Structure and Functioning of Estuarine and Nearshore EcosystemsnajwamohamadNo ratings yet
- TUGAS 08 Pengetahuan Fisik Laut'Document6 pagesTUGAS 08 Pengetahuan Fisik Laut'Rizaldi CaesarNo ratings yet
- Diversity: Macroinvertebrate Taxonomic Richness in Minimally Disturbed Streams On The Southeastern USA Coastal PlainDocument17 pagesDiversity: Macroinvertebrate Taxonomic Richness in Minimally Disturbed Streams On The Southeastern USA Coastal PlainViviana BravoNo ratings yet
- Marine Inverteb-WPS OfficeDocument22 pagesMarine Inverteb-WPS OfficembojefadhilNo ratings yet
- Fortes Et Al 2013Document12 pagesFortes Et Al 2013monteironetocassNo ratings yet
- Floodplain Wetland Biota in the Murray-Darling Basin: Water and Habitat RequirementsFrom EverandFloodplain Wetland Biota in the Murray-Darling Basin: Water and Habitat RequirementsKerrylee RogersNo ratings yet
- Shallow Groundwater Quality of Lahore City Along The River Ravi by Afaf AyeshaDocument9 pagesShallow Groundwater Quality of Lahore City Along The River Ravi by Afaf AyeshaShumaila MajeedNo ratings yet
- Assignment 1 Disaster& Risk Management Submitted By: Ezza Ahmad Butt Submitted To: Ma'Am Shumaila Submission Date: 24 SPET 2021 Semester 7Document5 pagesAssignment 1 Disaster& Risk Management Submitted By: Ezza Ahmad Butt Submitted To: Ma'Am Shumaila Submission Date: 24 SPET 2021 Semester 7Shumaila MajeedNo ratings yet
- 1 Groundwater Levels, Temperature and QualityDocument14 pages1 Groundwater Levels, Temperature and QualityShumaila MajeedNo ratings yet
- Droughts: Types of DroughtDocument3 pagesDroughts: Types of DroughtShumaila MajeedNo ratings yet
- Metropolitian Drain of Lahore CityDocument15 pagesMetropolitian Drain of Lahore CityShumaila MajeedNo ratings yet
- DLL January 9, 2024Document4 pagesDLL January 9, 2024Jezreel RabaNo ratings yet
- Narrative PHT3Document3 pagesNarrative PHT3Jericho Basmayor SajulanNo ratings yet
- Indiv Scores April24th & 25th 2010Document4 pagesIndiv Scores April24th & 25th 2010ABESC150No ratings yet
- Survive Mola Mola - OrvDocument35 pagesSurvive Mola Mola - OrvPhương Lương MaiNo ratings yet
- Aqua History PhilDocument27 pagesAqua History Philvirgil mae taghapNo ratings yet
- 4th Science Notes The HopeDocument67 pages4th Science Notes The Hopemasterm9800No ratings yet
- Calantas, Jana Karla C. BS Fisheries III: Land-Based Hatchery and Juvenile Production FarmDocument3 pagesCalantas, Jana Karla C. BS Fisheries III: Land-Based Hatchery and Juvenile Production FarmJana Karla CalantasNo ratings yet
- Catfish Farming Information Guide - AgrifarmingDocument5 pagesCatfish Farming Information Guide - AgrifarmingamolkajaleNo ratings yet
- Fisheries BiologyDocument29 pagesFisheries Biologybikashh100% (3)
- Theodoropoulou - Tuna Fishing in The Aegean-2013Document1 pageTheodoropoulou - Tuna Fishing in The Aegean-2013TeSoTrasNo ratings yet
- Chilled and Frozen Raw Fish PDFDocument26 pagesChilled and Frozen Raw Fish PDFMourad AttouchiNo ratings yet
- AquariumDocument65 pagesAquariumDaisy100% (4)
- PBS Baits E - Book Version 1Document48 pagesPBS Baits E - Book Version 1Dirk Henn0% (1)
- African Catfish (Clarias Gariepinus) : Introduction To African Catfish Hatchery Set Up and ManagementDocument14 pagesAfrican Catfish (Clarias Gariepinus) : Introduction To African Catfish Hatchery Set Up and ManagementHaymanot yenesewNo ratings yet
- International Standard Statistical Classification of Fishery Vessels by Vessel Types (ISSCFV, Rev.1, 2019)Document3 pagesInternational Standard Statistical Classification of Fishery Vessels by Vessel Types (ISSCFV, Rev.1, 2019)okta bedjoNo ratings yet
- What Is The Scope of Aquaculture?Document2 pagesWhat Is The Scope of Aquaculture?Louie YamonNo ratings yet
- Proximate Composition of Some Commercial Fish Feeds Sold in NigeriaDocument5 pagesProximate Composition of Some Commercial Fish Feeds Sold in NigeriaDR Ebthal GaberNo ratings yet
- Workshop On Assessment and Management of The Offshore Resources of South and Southeast AsiaDocument44 pagesWorkshop On Assessment and Management of The Offshore Resources of South and Southeast AsiadutvaNo ratings yet
- Optimalisasi Pemanfaatan Dan Pengelolaan Pangkalan Pendaratan Ikan (Ppi) Klaligi Kota Sorong Propinsi Papua BaratDocument150 pagesOptimalisasi Pemanfaatan Dan Pengelolaan Pangkalan Pendaratan Ikan (Ppi) Klaligi Kota Sorong Propinsi Papua BaratMuhammad IqbalNo ratings yet
- Use of Azolla Pinnata (RRL)Document9 pagesUse of Azolla Pinnata (RRL)Jennie Marlow AddisonNo ratings yet
- World Kids Colouring Day Magazine 2016Document12 pagesWorld Kids Colouring Day Magazine 2016Carmen DraghiaNo ratings yet
- 6120838371344Document6 pages6120838371344DeeNo ratings yet
- Fish Processing: Santiago CampusDocument24 pagesFish Processing: Santiago CampusRomnick Elgamo GalantoNo ratings yet
- Endemic Fish of The Birds Head - G. AllenDocument5 pagesEndemic Fish of The Birds Head - G. AllenRudy PriatnaNo ratings yet
- Learning Activity Sheet No.1 in Science 2: Key ConceptDocument25 pagesLearning Activity Sheet No.1 in Science 2: Key ConceptMary Grace BaldozaNo ratings yet
- Fish and Shellfish IdentificationDocument107 pagesFish and Shellfish IdentificationthewurzelNo ratings yet
- +1 Bio Zoo em Vol 1Document30 pages+1 Bio Zoo em Vol 1gowthamgokul206No ratings yet
- Elementary Group Pop-Quiz 115 ReadingDocument3 pagesElementary Group Pop-Quiz 115 ReadingChristophe Kodjo CharlecNo ratings yet
- Fish Species NotebookDocument20 pagesFish Species NotebookLead HeadNo ratings yet
- 2nd Quarter Assessment in Cookery 10Document3 pages2nd Quarter Assessment in Cookery 10Irish Borga100% (1)