Download as pdf or txt
You might also like
- Cambridge IGCSE Mathematics Core and Extended Workbook by Ric Pimentel, Terry Wall PDFDocument97 pagesCambridge IGCSE Mathematics Core and Extended Workbook by Ric Pimentel, Terry Wall PDFPrince Yug86% (22)
- Teachings in The Science of Osteopathy PDFDocument336 pagesTeachings in The Science of Osteopathy PDFAlex92% (12)
- List of Pharmaceutical Companies in Ahmedabad WithDocument22 pagesList of Pharmaceutical Companies in Ahmedabad Withsandipsc88% (8)
- Exam C Manual PDFDocument2 pagesExam C Manual PDFJohnathan0% (1)
- Manual Skoda Octavia 1,4 55kWDocument120 pagesManual Skoda Octavia 1,4 55kWCornea Horatiu Sebastian50% (2)
- Speech & Language Therapy in Practice, Spring 2002Document32 pagesSpeech & Language Therapy in Practice, Spring 2002Speech & Language Therapy in PracticeNo ratings yet
- Aspects of Urea Metabolism in Ruminants With Referen 1980 Journal of Dairy SDocument22 pagesAspects of Urea Metabolism in Ruminants With Referen 1980 Journal of Dairy SCtNo ratings yet
- 1 s2.0 S0022030275846972 MainDocument21 pages1 s2.0 S0022030275846972 MainAjoNo ratings yet
- Amino Acid Catabolism1Document5 pagesAmino Acid Catabolism1rolwinNo ratings yet
- Captulo 19 BaynesDocument15 pagesCaptulo 19 BaynesMaria Jose Monsalve CuencaNo ratings yet
- 02 Protein DigestionDocument44 pages02 Protein DigestionJohn MusaNo ratings yet
- Kuliah Fisum 8 Dan 9Document42 pagesKuliah Fisum 8 Dan 9Alloy Eginta GintingNo ratings yet
- Pengaruh Pemberian Tepung Gaplek - Urea Yang Dikukus Terhadap Konsumsi Dan Kecernaan Protein Serta Neraca Nitrogen Pada DombaDocument5 pagesPengaruh Pemberian Tepung Gaplek - Urea Yang Dikukus Terhadap Konsumsi Dan Kecernaan Protein Serta Neraca Nitrogen Pada DombaKikyNo ratings yet
- B. Katabolisme Asam Amino-1Document19 pagesB. Katabolisme Asam Amino-1M Sifal MaulanaNo ratings yet
- A.A MetabolismDocument31 pagesA.A Metabolismalexmichael2665No ratings yet
- 4 ProteinDocument11 pages4 ProteinconcepcionsofiakarylleNo ratings yet
- Ruminant Protein Nutrition: Lebih Di Kenal Dengan Metabolisme Nitrogen Dalam RumenDocument24 pagesRuminant Protein Nutrition: Lebih Di Kenal Dengan Metabolisme Nitrogen Dalam RumenIsnainiwnNo ratings yet
- Ijms 20 01587Document20 pagesIjms 20 01587LouiseNo ratings yet
- Orotic Acid Excretion and Arginine MetabolismDocument6 pagesOrotic Acid Excretion and Arginine MetabolismKevin Hernandez PerezNo ratings yet
- Immunonutrition: The Role of ArginineDocument7 pagesImmunonutrition: The Role of ArginineNatália LopesNo ratings yet
- Absortion AaDocument30 pagesAbsortion Aaandres herreraNo ratings yet
- C10 Protein and Amino Acid MetabolismDocument8 pagesC10 Protein and Amino Acid MetabolismSoraya D. Al-ObinayNo ratings yet
- Write Up 2Document8 pagesWrite Up 2Natanga GodfreyNo ratings yet
- Maiuolo 2016Document7 pagesMaiuolo 2016okaNo ratings yet
- Proteins and Protein MetabolismDocument9 pagesProteins and Protein MetabolismclaireNo ratings yet
- Biochem2-Week 7-Fates of N Group-Urea CycleDocument30 pagesBiochem2-Week 7-Fates of N Group-Urea CycleBùi Ngọc MaiNo ratings yet
- 6 Proteins Their Digestion & AbsorptionsDocument18 pages6 Proteins Their Digestion & AbsorptionshodaNo ratings yet
- BIOSINTESIS PROTlanjutanDocument31 pagesBIOSINTESIS PROTlanjutanAprilikkaearlyNo ratings yet
- Proteins and Amino AcidsDocument9 pagesProteins and Amino AcidsNatanga GodfreyNo ratings yet
- Watford 2003Document9 pagesWatford 2003josetuiran992No ratings yet
- Optigen Research Paper2Document18 pagesOptigen Research Paper2SUBRATA PARAINo ratings yet
- Development of Urea Products As Rumen Slow-ReleaseDocument11 pagesDevelopment of Urea Products As Rumen Slow-ReleaseEdtyva monichaNo ratings yet
- Digestion & Absorption ProteinsDocument44 pagesDigestion & Absorption ProteinsPutri Indah NirmalasariNo ratings yet
- 11 เมแทบอลิซึมของโปรตีนDocument44 pages11 เมแทบอลิซึมของโปรตีน22vk6svnb4No ratings yet
- Lectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-201-231Document31 pagesLectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-201-231Ken M'voulaboloNo ratings yet
- WameedMUCLecture_2021_92920280Document15 pagesWameedMUCLecture_2021_92920280odhiambovickniceNo ratings yet
- Regulation of Uric Acid Metabolism and Excretion PDFDocument7 pagesRegulation of Uric Acid Metabolism and Excretion PDFYulanda YomemotoNo ratings yet
- Nutritional and Genetical Factors in Uencing Nitrogen Metabolism and Excretion in Dairy Cows: A ReviewDocument9 pagesNutritional and Genetical Factors in Uencing Nitrogen Metabolism and Excretion in Dairy Cows: A ReviewIgor BaltaNo ratings yet
- ProteinsDocument22 pagesProteinsJeswanth NayakNo ratings yet
- Dosis UreaDocument12 pagesDosis UreaEdtyva monichaNo ratings yet
- Chemical Pathology BS-MLT 5Th SemesterDocument36 pagesChemical Pathology BS-MLT 5Th SemesterMuhammad AbdullahNo ratings yet
- Protein MetaboilisamDocument18 pagesProtein MetaboilisamSumit PandyaNo ratings yet
- Amino Acid MetabolismDocument15 pagesAmino Acid Metabolismshanto.tn98No ratings yet
- Ammonia and Urea CycleDocument17 pagesAmmonia and Urea CycleAboubakar Moalim Mahad moh'dNo ratings yet
- Catabolism of Amino Acids: DR - Ula Abbas ZekiDocument49 pagesCatabolism of Amino Acids: DR - Ula Abbas ZekinameNo ratings yet
- Topic: Protein Metabolism: Biochemistry & Biophysics: Paper-V, Unit-9Document14 pagesTopic: Protein Metabolism: Biochemistry & Biophysics: Paper-V, Unit-9aayushi tejwaniNo ratings yet
- Nutrient Dynamics in Aquaponic SystemsDocument3 pagesNutrient Dynamics in Aquaponic SystemsHenk StanderNo ratings yet
- Mariotti, AJCN, 2013Document8 pagesMariotti, AJCN, 2013Dianne Faye ManabatNo ratings yet
- Biochemistry II Protein (Metabolism of Amino Acids) (New Edition)Document78 pagesBiochemistry II Protein (Metabolism of Amino Acids) (New Edition)Rekti Viony100% (1)
- Chapter 15. Protein and Amino Acid MetabolismDocument42 pagesChapter 15. Protein and Amino Acid Metabolismrahul.kumar4504No ratings yet
- METABOLISME ASAM AMINO Protein BiologiDocument31 pagesMETABOLISME ASAM AMINO Protein BiologiAxzchiuu :vNo ratings yet
- Presentasi AmphibiaDocument30 pagesPresentasi Amphibiabima satria moektiNo ratings yet
- Methods To Study Degradation of Ruminant FeedsDocument15 pagesMethods To Study Degradation of Ruminant FeedsJ Jesus Bustamante GroNo ratings yet
- Urea and Ammonia Metabolism and The Control of Renal Nitrogen ExcretionDocument15 pagesUrea and Ammonia Metabolism and The Control of Renal Nitrogen ExcretionJames Cojab SacalNo ratings yet
- Effect of Ammonia Concentration On Rumen Microbial Protein Production in VitroDocument10 pagesEffect of Ammonia Concentration On Rumen Microbial Protein Production in VitroHidayat Anjar AsmaraniNo ratings yet
- Protein and Amino Acid MetabolismDocument32 pagesProtein and Amino Acid MetabolismVirag0% (1)
- Prot Eval Meth and Syst Ruminants OverviewDocument5 pagesProt Eval Meth and Syst Ruminants Overviewjss_bustamanteNo ratings yet
- My Answer 2Document3 pagesMy Answer 2Prince MoshehNo ratings yet
- 9a. Metabolisme Asam AminoDocument62 pages9a. Metabolisme Asam AminohimawarumNo ratings yet
- Protein, Nitrogen Katabolisme Dan Siklus UreaDocument35 pagesProtein, Nitrogen Katabolisme Dan Siklus UreaAnonymous QCMhA4wNgBNo ratings yet
- Amino Acid MetabolismDocument29 pagesAmino Acid MetabolismERIAS TENYWANo ratings yet
- Amino Acids Metabolism Final For Pharm 2014Document57 pagesAmino Acids Metabolism Final For Pharm 2014Getu LuchesaNo ratings yet
- Amino Acid Protein and Nucleic Acid Metabolism 20182019 LectureDocument18 pagesAmino Acid Protein and Nucleic Acid Metabolism 20182019 LectureMwanja MosesNo ratings yet
- Anaerobe: E. Gozde Ozbayram, Sabine Kleinsteuber, Marcell Nikolausz, Bahar Ince, Orhan InceDocument9 pagesAnaerobe: E. Gozde Ozbayram, Sabine Kleinsteuber, Marcell Nikolausz, Bahar Ince, Orhan InceSivakumarNo ratings yet
- A Compilation of Ligno-Cellulose Feedstock and Related Research for Feed, Food and EnergyFrom EverandA Compilation of Ligno-Cellulose Feedstock and Related Research for Feed, Food and EnergyNo ratings yet
- On Digestive Proteolysis: Being the Cartwright Lectures for 1894From EverandOn Digestive Proteolysis: Being the Cartwright Lectures for 1894No ratings yet
- Cambridge International General Certificate of Secondary EducationDocument3 pagesCambridge International General Certificate of Secondary EducationkabirNo ratings yet
- Manitou Telehandler MLT 840 MLT 1040 Ps JD St4 s1 Genuine Parts Catalogue 647541en 09 2015Document22 pagesManitou Telehandler MLT 840 MLT 1040 Ps JD St4 s1 Genuine Parts Catalogue 647541en 09 2015mscassandravasquez120985pgc100% (106)
- Fama FraternitatisDocument10 pagesFama Fraternitatistom watersNo ratings yet
- Orientation Special... : Byu Indian Education Department SUMMER 1979Document4 pagesOrientation Special... : Byu Indian Education Department SUMMER 1979blopbookNo ratings yet
- Tool 20-05-21Document13 pagesTool 20-05-21naga maniNo ratings yet
- Chapter 6 Consumption and SavingsDocument1 pageChapter 6 Consumption and SavingsEnter any name or emailNo ratings yet
- Eng Taas-08 Talmud Eser SefirotDocument63 pagesEng Taas-08 Talmud Eser Sefirotne0h_16_196547097No ratings yet
- Schedule Waste HandlingDocument3 pagesSchedule Waste Handlingosha911No ratings yet
- Https Sis Cat Com Sisweb Sisweb Techdoc Techdoc Print Page 2Document4 pagesHttps Sis Cat Com Sisweb Sisweb Techdoc Techdoc Print Page 2Tshidiso LekalakalaNo ratings yet
- Table 60001 Concrete DurabilityDocument1 pageTable 60001 Concrete DurabilityZaid JamilNo ratings yet
- EPC Saur Energy InternationalDocument3 pagesEPC Saur Energy InternationalNarendra PatelNo ratings yet
- Licencia KmsDocument2 pagesLicencia Kmseliecer david solanoNo ratings yet
- Invinsense OMDRDocument7 pagesInvinsense OMDRinfoperceptNo ratings yet
- Harpa Concert Hall & Conference CenterDocument18 pagesHarpa Concert Hall & Conference Centershenaz AfroseNo ratings yet
- Inventarisasi Pohon Untuk Pengelolaan Pohon Asuh Di Hutan Adat Rantau KermasDocument11 pagesInventarisasi Pohon Untuk Pengelolaan Pohon Asuh Di Hutan Adat Rantau KermasAndreas WahyuNo ratings yet
- CarbonCure Whitepaper Impact of CO2 Utilization in Fresh Concrete On Corrosion of Steel ReinforcementDocument6 pagesCarbonCure Whitepaper Impact of CO2 Utilization in Fresh Concrete On Corrosion of Steel ReinforcementSakineNo ratings yet
- Rough Surface Scattering ModelsDocument11 pagesRough Surface Scattering ModelshakimNo ratings yet
- Today1 AchievementTestUnits3 4extensionDocument2 pagesToday1 AchievementTestUnits3 4extensionEmii GarcíaNo ratings yet
- Science 9 Q4 Intro To KinematicsDocument54 pagesScience 9 Q4 Intro To KinematicsSire WillNo ratings yet
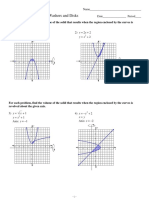
- 07 - Volume Washers and DisksDocument4 pages07 - Volume Washers and DisksDhatchana SubramaniamNo ratings yet
- Multikolinieritas: Regression Analysis: X1 Versus X2, X3Document8 pagesMultikolinieritas: Regression Analysis: X1 Versus X2, X3ine_bz1530No ratings yet
- AWS Welding Standard PreviewDocument5 pagesAWS Welding Standard PreviewRavi YadavNo ratings yet
- Bronchoscopy: Pedia Ward 1 GroupDocument19 pagesBronchoscopy: Pedia Ward 1 GroupMARIANN JEAN ANDREA CULANAG MATALINESNo ratings yet
- Laboratory or Rabbit or Pet Animal Productions PDFDocument19 pagesLaboratory or Rabbit or Pet Animal Productions PDFPublic InterestNo ratings yet