Download as pdf or txt
You might also like
- NOAA Shark Identification PlacardsDocument6 pagesNOAA Shark Identification PlacardsMS Dept of Marine Resources100% (2)
- Discoveries of New Mammal SpeciesDocument7 pagesDiscoveries of New Mammal Species--No ratings yet
- Abba e Tal 2012-MammaliaDocument15 pagesAbba e Tal 2012-MammaliaEkaterina SavchenkoNo ratings yet
- Good Man 2015Document9 pagesGood Man 2015PochoNo ratings yet
- Bradypus Journal of Mammalian Evolution-2019Document24 pagesBradypus Journal of Mammalian Evolution-2019Manuel RuizNo ratings yet
- Usnea Oreophila (Parmeliaceae), A New Saxicolous Species From The Mountains of BrazilDocument9 pagesUsnea Oreophila (Parmeliaceae), A New Saxicolous Species From The Mountains of BrazilRafael Felipe de AlmeidaNo ratings yet
- A Review of the Middle American Tree Frogs of the Genus PtychohylaFrom EverandA Review of the Middle American Tree Frogs of the Genus PtychohylaNo ratings yet
- Artificial Keys To The Genera of Living Stalked CRDocument33 pagesArtificial Keys To The Genera of Living Stalked CRAurelia Izzatul AzkaNo ratings yet
- Clarc Usena 2016Document10 pagesClarc Usena 2016francibelkNo ratings yet
- 2017 Waichert Et Al More Records PompilidaeDocument7 pages2017 Waichert Et Al More Records PompilidaeZhary VanegasNo ratings yet
- Tadpole, Oophagy, Advertisement Call, and Geographic Distribution of Aparasphenodon Arapapa Pimenta, Napoli and Haddad 2009 (Anura, Hylidae)Document6 pagesTadpole, Oophagy, Advertisement Call, and Geographic Distribution of Aparasphenodon Arapapa Pimenta, Napoli and Haddad 2009 (Anura, Hylidae)Guilherme SousaNo ratings yet
- Barasoainetal 2019Document19 pagesBarasoainetal 2019Huáscar Bustillos CayojaNo ratings yet
- Nesting Biology of A New High Andean Bee, Anthophora Walteri Gonzalez (Hymenoptera: Apidae: Anthophorini)Document10 pagesNesting Biology of A New High Andean Bee, Anthophora Walteri Gonzalez (Hymenoptera: Apidae: Anthophorini)Pablo AndresNo ratings yet
- The Geographic Pattern of Distribution of The Genus - Em-RhopaluruDocument16 pagesThe Geographic Pattern of Distribution of The Genus - Em-RhopaluruAlbert Antonio Flores MejiaNo ratings yet
- Gonzalez2014 12247 LiphanthusDocument13 pagesGonzalez2014 12247 Liphanthusyeison calizaya meloNo ratings yet
- B. Chucantiensis2 Garcs Miranda Fuentes Batista December 2016 PDFDocument114 pagesB. Chucantiensis2 Garcs Miranda Fuentes Batista December 2016 PDFAgroecologia UTNo ratings yet
- A New Species of Petracola (Squamata: Gymnophthalmidae) From Río Abiseo National ParkDocument8 pagesA New Species of Petracola (Squamata: Gymnophthalmidae) From Río Abiseo National ParkGiuseppe GagliardiNo ratings yet
- Handbook of The Mammals of The World 3 PrimatesDocument3 pagesHandbook of The Mammals of The World 3 Primatesltorresm6No ratings yet
- 348 1192 1 SMDocument90 pages348 1192 1 SMmarcio vieiraNo ratings yet
- Notes On Natural History and Call Description of The Critically Endangered Plectrohyla Avia (Anura: Hylidae) From Chiapas, MexicoDocument7 pagesNotes On Natural History and Call Description of The Critically Endangered Plectrohyla Avia (Anura: Hylidae) From Chiapas, Mexicoangy kudoNo ratings yet
- Torres-Carvajal 2007 StenocercusDocument104 pagesTorres-Carvajal 2007 StenocercusArenita PerdidaNo ratings yet
- Rhinella Major Rhinella GR Margaritifera Food For SpidersDocument7 pagesRhinella Major Rhinella GR Margaritifera Food For SpidersPatrick SanchesNo ratings yet
- Body Mass Estimation in Lujanian (Late Pleistocene-Early Holocene of South America) Mammal MegafaunaDocument23 pagesBody Mass Estimation in Lujanian (Late Pleistocene-Early Holocene of South America) Mammal MegafaunaVivi LiaoNo ratings yet
- American J of Botany - 2004 - Fritsch - Phylogeny and Circumscription of The Near Endemic Brazilian Tribe MicrolicieaeDocument10 pagesAmerican J of Botany - 2004 - Fritsch - Phylogeny and Circumscription of The Near Endemic Brazilian Tribe Microlicieaemateus silvaNo ratings yet
- Vitt & Vangilder (1983) - Ecology of Snake Community in The Northeastern BrazilDocument24 pagesVitt & Vangilder (1983) - Ecology of Snake Community in The Northeastern BrazilEd MyersNo ratings yet
- Systematics and Evolution of Spiders (Araneae) : Jonathan A. CoddingtonDocument28 pagesSystematics and Evolution of Spiders (Araneae) : Jonathan A. CoddingtonHernanFigueredoNo ratings yet
- Revision of The Genus Murunducaris Copepoda HarpacDocument22 pagesRevision of The Genus Murunducaris Copepoda HarpacpcorgoNo ratings yet
- A Review of the Middle American Tree Frogs of the Genus PtychohylaFrom EverandA Review of the Middle American Tree Frogs of the Genus PtychohylaNo ratings yet
- First Record of Centruroides Edwardsii (Gervais, 1843) and New Geographic Distribution of Tityus Ocelote Francke - Stockwell, 1987 (Scorpiones - Buthidae) in PanamáDocument6 pagesFirst Record of Centruroides Edwardsii (Gervais, 1843) and New Geographic Distribution of Tityus Ocelote Francke - Stockwell, 1987 (Scorpiones - Buthidae) in PanamáCarlos MoránNo ratings yet
- Philodryas Patagoniensis - Diet - Lopez (2003)Document2 pagesPhilodryas Patagoniensis - Diet - Lopez (2003)Danilo CapelaNo ratings yet
- International Journal Primatology Definitive 10.1007Document39 pagesInternational Journal Primatology Definitive 10.1007Manuel RuizNo ratings yet
- Eugerres MexicanusDocument12 pagesEugerres MexicanusFlavioNo ratings yet
- ENCISO CALLE 2017 Rediscovery of Atelopus Subornatus WernerDocument4 pagesENCISO CALLE 2017 Rediscovery of Atelopus Subornatus WernerRaiza Nathaly Castañeda BonillaNo ratings yet
- A New Species of The Genus Stenocercus (Iguania Tropiduridae) From The Peruvian Pacific Coast (Ica Region) PDFDocument15 pagesA New Species of The Genus Stenocercus (Iguania Tropiduridae) From The Peruvian Pacific Coast (Ica Region) PDFRenzo Alessandro Pacherres NietoNo ratings yet
- The Genus Triturus: 612 Book ReviewsDocument2 pagesThe Genus Triturus: 612 Book Reviewsdan_cogalniceanu7989No ratings yet
- Supérvivencia y LogebidadDocument10 pagesSupérvivencia y LogebidadDaniel VargasNo ratings yet
- Riva 1996Document9 pagesRiva 1996erikaNo ratings yet
- The Sixth Extinction CrisisDocument17 pagesThe Sixth Extinction CrisisJose Manuel Lara PeñaNo ratings yet
- 04 Voilloud Colombian Peruvian AmazonDocument19 pages04 Voilloud Colombian Peruvian AmazonBibiana DuarteNo ratings yet
- A New Species of Burrowing CrayfishDocument13 pagesA New Species of Burrowing CrayfishDavid Orlando Carmona LagosNo ratings yet
- Molecular Phylogenetics and EvolutionDocument8 pagesMolecular Phylogenetics and EvolutionHuáscar Bustillos CayojaNo ratings yet
- PristimantisDocument22 pagesPristimantisArenita PerdidaNo ratings yet
- Armenteros 2009 TerschellingiaDocument11 pagesArmenteros 2009 Terschellingiaalex_manoNo ratings yet
- Two New Sympatric Species of Stenocercus (Squamata Iguania) From The Interandean Valley of The Mantaro River, Peru PDFDocument21 pagesTwo New Sympatric Species of Stenocercus (Squamata Iguania) From The Interandean Valley of The Mantaro River, Peru PDFRenzo Alessandro Pacherres NietoNo ratings yet
- Nothopsis Snake NoteDocument2 pagesNothopsis Snake NoteArco Iris CallejaNo ratings yet
- Acevedo 2016Document14 pagesAcevedo 2016parra tvNo ratings yet
- Reproductive Biology of A Bromeligenous Frog Endemic To The Atlantic ForestDocument14 pagesReproductive Biology of A Bromeligenous Frog Endemic To The Atlantic ForestamandasantiagoNo ratings yet
- 1 FloresVillela1990Document3 pages1 FloresVillela1990Livia PereiraNo ratings yet
- Neo Elm Is GuaraniDocument14 pagesNeo Elm Is GuaraniCarlos Llanos VasquezNo ratings yet
- Herpetological ReviewDocument61 pagesHerpetological ReviewbdsbioNo ratings yet
- A New Species of Hsunycteris From Northeastern Peru - Velazco Et Al. 2017Document29 pagesA New Species of Hsunycteris From Northeastern Peru - Velazco Et Al. 2017garciafranger97No ratings yet
- Cheracebus Aquinoi N SP (MS 2022)Document16 pagesCheracebus Aquinoi N SP (MS 2022)Walter VegaNo ratings yet
- Lamonicaetal 2007Document7 pagesLamonicaetal 2007Oscar Daniel Medina BarriosNo ratings yet
- Phylogeny and Biogeography of Asian Pit VipersDocument32 pagesPhylogeny and Biogeography of Asian Pit VipersHaydee ParasNo ratings yet
- Cryptotriton 2015Document18 pagesCryptotriton 2015FLOR DE MARIA MORALES ARROYONo ratings yet
- CalliopsisDocument1 pageCalliopsisJS At InferosNo ratings yet
- El Genero Astrocaryum ArecaceaeDocument19 pagesEl Genero Astrocaryum ArecaceaeJorge Luis Contreras HerreraNo ratings yet
- Biodiversity Web1 Diversity2021Document40 pagesBiodiversity Web1 Diversity2021GarryNo ratings yet
- Perbiche Nevesetal2015Document111 pagesPerbiche Nevesetal2015PriscilaNo ratings yet
- Morphological Characterization in The Genus PassifloraDocument29 pagesMorphological Characterization in The Genus PassifloraGOD EXONo ratings yet
- Rovito (2010) Bolitoglossa ZacapensisDocument12 pagesRovito (2010) Bolitoglossa ZacapensisFLOR DE MARIA MORALES ARROYONo ratings yet
- Key To Genera of Larvae of Australian Chironomidae (Diptera)Document32 pagesKey To Genera of Larvae of Australian Chironomidae (Diptera)Carli RodríguezNo ratings yet
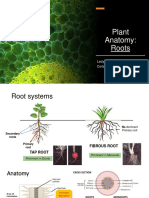
- Plant Anatomy RootsDocument6 pagesPlant Anatomy RootsCarli RodríguezNo ratings yet
- Trichoptera Phylogeny Adaptations and BiogeographyDocument13 pagesTrichoptera Phylogeny Adaptations and BiogeographyCarli RodríguezNo ratings yet
- Trichoptera Phylogeny Adaptations and BiogeographyDocument13 pagesTrichoptera Phylogeny Adaptations and BiogeographyCarli RodríguezNo ratings yet
- Merrit & Cummins Grupos Funcionales AlimenticiosDocument77 pagesMerrit & Cummins Grupos Funcionales AlimenticiosCarli Rodríguez100% (1)
- Birds Small Animals 2020-2021Document170 pagesBirds Small Animals 2020-2021Talia GodoiNo ratings yet
- Vertebrate Evolution: Classis AmphibiaDocument18 pagesVertebrate Evolution: Classis AmphibiaAghnia ChoiNo ratings yet
- Development of Face & Oral CavityDocument56 pagesDevelopment of Face & Oral CavityGanesh KulkarniNo ratings yet
- Story Telling NewDocument4 pagesStory Telling NewSMPYPIDARUSSALAM1No ratings yet
- Organic Chemistry 8Th Edition Bruice Test Bank Full Chapter PDFDocument67 pagesOrganic Chemistry 8Th Edition Bruice Test Bank Full Chapter PDFjunemojarrazqmxj100% (7)
- SOAL Kls 9 GENAP 2024Document2 pagesSOAL Kls 9 GENAP 2024yammiterritory338No ratings yet
- ĐỌC HIỂU - TEST 15Document3 pagesĐỌC HIỂU - TEST 15Giang GiangNo ratings yet
- Question Paper Exploring Effects and ImpactDocument1 pageQuestion Paper Exploring Effects and ImpactN RamNo ratings yet
- Chapter 2Document8 pagesChapter 2Mayank AggarwalNo ratings yet
- Anatomical Landmarks - PCPDocument14 pagesAnatomical Landmarks - PCPSara Sultana0% (1)
- CockroachDocument5 pagesCockroachdeveshjayakumaryadavNo ratings yet
- Preschool - MayDocument1 pagePreschool - MayDanijela KrunićNo ratings yet
- Animal KingdomDocument13 pagesAnimal KingdomAngel MaryNo ratings yet
- SZL 104 Practical 2 Phylum AnnelidaDocument5 pagesSZL 104 Practical 2 Phylum AnnelidaCheries RhodaNo ratings yet
- Vogelwarte 56 2018-3 DO-G Artenliste DEDocument54 pagesVogelwarte 56 2018-3 DO-G Artenliste DEGAHANo ratings yet
- Weekly Science P4 February 2021 PDFDocument4 pagesWeekly Science P4 February 2021 PDFMs SinarNo ratings yet
- Osteichthyes PDFDocument50 pagesOsteichthyes PDFmuhammadismailNo ratings yet
- Sowerby - Tankerville CatalogueDocument166 pagesSowerby - Tankerville CatalogueDavid StrongNo ratings yet
- Christensen and Winter 1997Document14 pagesChristensen and Winter 1997Soledad VictoriaNo ratings yet
- Week 8 Phylum PlatyhelminthesDocument29 pagesWeek 8 Phylum PlatyhelminthesAlyssa Jane AbellonNo ratings yet
- Untitled 2Document6 pagesUntitled 2Dr Prashant ChauhanNo ratings yet
- BENTHOS (Benthic Animals)Document7 pagesBENTHOS (Benthic Animals)Christine Israel AbrahamNo ratings yet
- 1apogonidae IvanceDocument40 pages1apogonidae IvanceCrimsondustNo ratings yet
- Which Vertebrate Has Two LivesDocument39 pagesWhich Vertebrate Has Two LivesgiselafariniaprofNo ratings yet
- JawsDocument2 pagesJawsbakircid0244No ratings yet
- Jolly Phonics Level 1 and 2 PracticeDocument133 pagesJolly Phonics Level 1 and 2 PracticeAnastasiia VoskNo ratings yet
- Druid - Wild Shape Beast ListDocument46 pagesDruid - Wild Shape Beast ListJan NuñezNo ratings yet
- Chapter 1: Introduction The Phylum Chordata: Big Four Protochordates HeadDocument14 pagesChapter 1: Introduction The Phylum Chordata: Big Four Protochordates HeadMacy MarianNo ratings yet
- រឿង ប្រវត្តិភ្នំសំពៅDocument20 pagesរឿង ប្រវត្តិភ្នំសំពៅTan Srey IngNo ratings yet