Download as pdf or txt
You might also like
- Santos Et Al 2021 (Tetra Olho de Fogo)Document5 pagesSantos Et Al 2021 (Tetra Olho de Fogo)Higo AbeNo ratings yet
- J Jinsphys 2010 06 015Document6 pagesJ Jinsphys 2010 06 015Belay AbdissaNo ratings yet
- Growth Performance of Angelfish PterophyDocument7 pagesGrowth Performance of Angelfish PterophyArdian RahmanNo ratings yet
- B5 5 Sanches 2011 Time and Temperature On The Storage of Oocytes From Jundi Catfish Rhamdia Quelen - En.idDocument6 pagesB5 5 Sanches 2011 Time and Temperature On The Storage of Oocytes From Jundi Catfish Rhamdia Quelen - En.idSilfia AgustinaNo ratings yet
- Are Lizards Feeling The Heat - A Tale of Ecology and Evolution Under Two TemperaturesDocument12 pagesAre Lizards Feeling The Heat - A Tale of Ecology and Evolution Under Two TemperatureschhoumioumaymaNo ratings yet
- Lood Values of Cortisol, Glucose, and Lactate in Healthy Green TurtleDocument7 pagesLood Values of Cortisol, Glucose, and Lactate in Healthy Green TurtleVitor FernandoNo ratings yet
- 1 s2.0 S2352340923008880 MainDocument6 pages1 s2.0 S2352340923008880 Maineng.cherllekallyNo ratings yet
- Fish StarvationDocument8 pagesFish Starvationdaniel cretuNo ratings yet
- Thesis 0.8Document9 pagesThesis 0.8jocyeoNo ratings yet
- Animals 12 03473Document17 pagesAnimals 12 03473Claudia Cristina Paro de PazNo ratings yet
- Project ThesisDocument29 pagesProject ThesisImmanuvelNo ratings yet
- The Effects of Larval Diet On Adult Life-History Traits of The Black Soldier Y, Hermetia Illucens (Diptera: Stratiomyidae)Document9 pagesThe Effects of Larval Diet On Adult Life-History Traits of The Black Soldier Y, Hermetia Illucens (Diptera: Stratiomyidae)Евгений ЯворскийNo ratings yet
- Full download Effect of protein on the post-ingestive response of tilapia fed encapsulated diets Leandro S. Costa file pdf all chapter on 2024Document32 pagesFull download Effect of protein on the post-ingestive response of tilapia fed encapsulated diets Leandro S. Costa file pdf all chapter on 2024aqasoyalardeNo ratings yet
- Effect of Protein On The Post Ingestive Response of Tilapia Fed Encapsulated Diets Leandro S Costa Full Chapter PDFDocument24 pagesEffect of Protein On The Post Ingestive Response of Tilapia Fed Encapsulated Diets Leandro S Costa Full Chapter PDFmrdendafydd100% (8)
- Bauer Et Al 2020 - Diet Moistire C MacellariaDocument10 pagesBauer Et Al 2020 - Diet Moistire C Macellariastephani.evelynNo ratings yet
- Effect of Temperature On Growth and Survival of Guppy (Poecilia Reticulata)Document7 pagesEffect of Temperature On Growth and Survival of Guppy (Poecilia Reticulata)Angie Lorena ParadaNo ratings yet
- Puvanendran 2013Document10 pagesPuvanendran 2013Belay AbdissaNo ratings yet
- La HomeopatíaDocument7 pagesLa HomeopatíaJonatan VelezNo ratings yet
- The Embryonic Development of Clarias Gariepinus Fertilized Eggs Subjected To Different Water Temperature Interval in An Indoor Hatchery in JosDocument6 pagesThe Embryonic Development of Clarias Gariepinus Fertilized Eggs Subjected To Different Water Temperature Interval in An Indoor Hatchery in Joswahyuni fanggi tasikNo ratings yet
- Zebrafish Assay Halili and QuilangBIOTA FinalDocument10 pagesZebrafish Assay Halili and QuilangBIOTA FinalGot SevenNo ratings yet
- Baldisserotop HDocument8 pagesBaldisserotop HJackson TrianaNo ratings yet
- Reproduccion Neritina Zebra PDFDocument8 pagesReproduccion Neritina Zebra PDFVictor PereaNo ratings yet
- Sfakianakis 2004Document18 pagesSfakianakis 2004Belay AbdissaNo ratings yet
- Fphys 14 1271809Document3 pagesFphys 14 1271809Zohar IbarraNo ratings yet
- Macrohirus) and Crappie (Pomoxis Anularis) at Varied Time Intervals andDocument13 pagesMacrohirus) and Crappie (Pomoxis Anularis) at Varied Time Intervals andkkkmiNo ratings yet
- Tropea Et Al 2012 PDFDocument8 pagesTropea Et Al 2012 PDFAndrea Gabriela RuizNo ratings yet
- Biotec Marina 2Document17 pagesBiotec Marina 2Piero PerezNo ratings yet
- Mini Review ArticleDocument7 pagesMini Review ArticleEsabel LlanoNo ratings yet
- Hormonal Manipulation of Fishes As An Aquacultural TechniqueDocument11 pagesHormonal Manipulation of Fishes As An Aquacultural TechniqueRommel CorpuzNo ratings yet
- 2018 Siqueire-Silve Biotechnology Applied To Fish ReproductionDocument14 pages2018 Siqueire-Silve Biotechnology Applied To Fish ReproductionloxibknNo ratings yet
- Physiological Responses of Gopher TortoisesDocument9 pagesPhysiological Responses of Gopher TortoisesVitor FernandoNo ratings yet
- Optimal Salinity and Temperature For Early Developmental Stages of Penaeus Merguiensis de ManDocument10 pagesOptimal Salinity and Temperature For Early Developmental Stages of Penaeus Merguiensis de ManOttoman CavalierNo ratings yet
- Ambient Temperature Influences Birds Decisions To Eat To 2013 Animal BehaviDocument8 pagesAmbient Temperature Influences Birds Decisions To Eat To 2013 Animal BehaviAnnie van BlaricumNo ratings yet
- Wu Et Al-2000-Entomologia Experimentalis Et Applicata ALIMENTAÇÃO TRICHOGRAMMADocument8 pagesWu Et Al-2000-Entomologia Experimentalis Et Applicata ALIMENTAÇÃO TRICHOGRAMMADagdaniNo ratings yet
- Nursery of Litopenaeus Vannamei Reared IDocument9 pagesNursery of Litopenaeus Vannamei Reared IM RAJASEKARANNo ratings yet
- Seasonal Variation of Energy Reserves and Reproduction in Neotropical Bats Molossus MolossusDocument7 pagesSeasonal Variation of Energy Reserves and Reproduction in Neotropical Bats Molossus MolossusÉrica MunhozNo ratings yet
- Early Ontogenesis of The Angelfish, Pterophyllum Scalare Schultze, 1823 (Cichlidae)Document10 pagesEarly Ontogenesis of The Angelfish, Pterophyllum Scalare Schultze, 1823 (Cichlidae)kusuma wardanaNo ratings yet
- Synergistic Effect of X-Ray Irradiation and Sodium Hypochlorite Against Salmonella Enterica Serovar Typhimurium Biofilms On Quail EggshellsDocument7 pagesSynergistic Effect of X-Ray Irradiation and Sodium Hypochlorite Against Salmonella Enterica Serovar Typhimurium Biofilms On Quail EggshellsCaro RestrepoNo ratings yet
- Ecophysiology of MammalsDocument17 pagesEcophysiology of MammalsYuliana AguileraNo ratings yet
- Pr. BiodoversitasDocument7 pagesPr. BiodoversitasdioNo ratings yet
- AnfíbiosDocument11 pagesAnfíbiosRita PinheiroNo ratings yet
- 1 s2.0 S0379073814004058 MainDocument6 pages1 s2.0 S0379073814004058 Mainstephani.evelynNo ratings yet
- Intestinal Parasite Infections and Fecal Steroid Levels in Wild ChimpanzeesDocument5 pagesIntestinal Parasite Infections and Fecal Steroid Levels in Wild ChimpanzeesDavi FragosoNo ratings yet
- Network Analysis of Gut Microbiota Literature: An Overview of The Research Landscape in Non-Human Animal StudiesDocument8 pagesNetwork Analysis of Gut Microbiota Literature: An Overview of The Research Landscape in Non-Human Animal Studiesvicrisyihab24No ratings yet
- Studies On The Reproductive Biology and Captive Breeding of Channa Aurantimaculata, An Endemic Fish From AssamDocument7 pagesStudies On The Reproductive Biology and Captive Breeding of Channa Aurantimaculata, An Endemic Fish From AssamChimo PissangNo ratings yet
- Bosmin ADocument4 pagesBosmin AÉrica OliveiraNo ratings yet
- Tempo and Mode of Genome Evolution in The Budding Yeast SubphylumDocument34 pagesTempo and Mode of Genome Evolution in The Budding Yeast SubphylumHARSH VARSHNEYNo ratings yet
- ResumejurnalDocument2 pagesResumejurnalabdul basithNo ratings yet
- Revista Brasileira de ZootecniaDocument7 pagesRevista Brasileira de ZootecniayutefupNo ratings yet
- Acute Effect of Cypermethrin On Nutrient Utilization and Histopathology of Clarias Gariepinus (Burchell, 1822)Document10 pagesAcute Effect of Cypermethrin On Nutrient Utilization and Histopathology of Clarias Gariepinus (Burchell, 1822)IJAR JOURNALNo ratings yet
- The Effect of Relative Humidity On Black Soldier y Development: A Study Using Two Dietary ResourcesDocument10 pagesThe Effect of Relative Humidity On Black Soldier y Development: A Study Using Two Dietary ResourcesMuhammad Sya FiqNo ratings yet
- 1 Lifestyles in Transition, Evolution and Natural History of Genus LactobacillusDocument22 pages1 Lifestyles in Transition, Evolution and Natural History of Genus Lactobacillusnalar patrisiaNo ratings yet
- Preprint Zarski 2019Document38 pagesPreprint Zarski 2019Alejandro Giraldo ObregónNo ratings yet
- PDF 529Document12 pagesPDF 529Alliah MendozaNo ratings yet
- Urata - Etal - Larval Development of The Oriental Lancelet - BranchiostomaDocument11 pagesUrata - Etal - Larval Development of The Oriental Lancelet - BranchiostomaRochyRiquelmeNo ratings yet
- Science and Justice: Lauren P. Chun, Marcus J. Miguel, Emily N. Junkins, Shari L. Forbes, David O. CarterDocument8 pagesScience and Justice: Lauren P. Chun, Marcus J. Miguel, Emily N. Junkins, Shari L. Forbes, David O. CarterMiguel NadalNo ratings yet
- Colston Et Al-2016-Molecular Ecology2222Document25 pagesColston Et Al-2016-Molecular Ecology2222Diana CalderaNo ratings yet
- Effects of Rotifer Feeding Frequency On Growth and Survival Rate of Early Larval Stages of Mud Crab, Scylla OlivaceaDocument11 pagesEffects of Rotifer Feeding Frequency On Growth and Survival Rate of Early Larval Stages of Mud Crab, Scylla Olivaceachris patrickNo ratings yet
- 1-s2.0-S003257911944827X-mainDocument5 pages1-s2.0-S003257911944827X-mainsanielNo ratings yet
- Success Factors for Fish Larval ProductionFrom EverandSuccess Factors for Fish Larval ProductionLuis ConceicaoNo ratings yet
- Effects of Temperature On Feeding and Digestive Processes in FishDocument15 pagesEffects of Temperature On Feeding and Digestive Processes in FishBelay AbdissaNo ratings yet
- Puvanendran 2013Document10 pagesPuvanendran 2013Belay AbdissaNo ratings yet
- J Jinsphys 2010 06 015Document6 pagesJ Jinsphys 2010 06 015Belay AbdissaNo ratings yet
- The Effect of Temperature and Fish Size On Growth and Feed Efficiency Ratio of Juvenile Spotted Wolffish Anarhichas MinorDocument16 pagesThe Effect of Temperature and Fish Size On Growth and Feed Efficiency Ratio of Juvenile Spotted Wolffish Anarhichas MinorBelay AbdissaNo ratings yet
- Morphological Diversity and Age DependenDocument17 pagesMorphological Diversity and Age DependenBelay AbdissaNo ratings yet
- The Influence of Temperature On The Development of Baltic Sea Sprat (Sprattus Sprattus) Eggs and Yolk Sac LarvaeDocument12 pagesThe Influence of Temperature On The Development of Baltic Sea Sprat (Sprattus Sprattus) Eggs and Yolk Sac LarvaeBelay AbdissaNo ratings yet
- Lahnsteiner 2012Document10 pagesLahnsteiner 2012Belay AbdissaNo ratings yet
- The Temperature-Size Rule Emerges From Ontogenetic Differences Between Growth and Development RatesDocument10 pagesThe Temperature-Size Rule Emerges From Ontogenetic Differences Between Growth and Development RatesBelay AbdissaNo ratings yet
- Effects of Hyperthyroidism On The: Labeobarbus ( Barbus) Intermedius (Cyprinidae) Early Larval Melanophores DevelopmentDocument4 pagesEffects of Hyperthyroidism On The: Labeobarbus ( Barbus) Intermedius (Cyprinidae) Early Larval Melanophores DevelopmentBelay AbdissaNo ratings yet
- Sfakianakis 2004Document18 pagesSfakianakis 2004Belay AbdissaNo ratings yet
- Rodrigues Galdino2009Document14 pagesRodrigues Galdino2009Belay AbdissaNo ratings yet
- Rice Fish Production Strategies in The CDocument6 pagesRice Fish Production Strategies in The CBelay AbdissaNo ratings yet
- s41598 021 88306 4Document11 pagess41598 021 88306 4Belay AbdissaNo ratings yet
- Marine Environmental Research: Marie Vagner, José-Luis Zambonino-Infante, David Mazurais TDocument20 pagesMarine Environmental Research: Marie Vagner, José-Luis Zambonino-Infante, David Mazurais TBelay AbdissaNo ratings yet
- Ontogenetic Patterns in Thermal Adaptation of Fish vs. Long-Term Temperature Trends in Large RiversDocument9 pagesOntogenetic Patterns in Thermal Adaptation of Fish vs. Long-Term Temperature Trends in Large RiversBelay AbdissaNo ratings yet
- Fishfarm 3 RdproofDocument38 pagesFishfarm 3 RdproofBelay AbdissaNo ratings yet
- Transactions of The American Fisheries SocietyDocument9 pagesTransactions of The American Fisheries SocietyBelay AbdissaNo ratings yet
- Georgakopoulou 2007Document14 pagesGeorgakopoulou 2007Belay AbdissaNo ratings yet
- Growth Performance of Clarias GariepinusDocument8 pagesGrowth Performance of Clarias GariepinusBelay AbdissaNo ratings yet
- Barulin 2Document10 pagesBarulin 2Belay AbdissaNo ratings yet
- Bolotovskiy 2011Document10 pagesBolotovskiy 2011Belay AbdissaNo ratings yet
- Transactions of The American Fisheries SocietyDocument8 pagesTransactions of The American Fisheries SocietyBelay AbdissaNo ratings yet
- Effect of Egg Incubation Temperature On The Occurrence of Skeletal Deformities in Solea SenegalensisDocument6 pagesEffect of Egg Incubation Temperature On The Occurrence of Skeletal Deformities in Solea SenegalensisBelay AbdissaNo ratings yet
- Sperm Cryopreservation of The Critically Endangered Olive Barb (Sarpunti) Puntius Sarana (Hamilton, 1822)Document8 pagesSperm Cryopreservation of The Critically Endangered Olive Barb (Sarpunti) Puntius Sarana (Hamilton, 1822)Belay AbdissaNo ratings yet
- Dicentrarchus Labrax: Temperature-Induced Ontogenetic Plasticity in Sea BassDocument14 pagesDicentrarchus Labrax: Temperature-Induced Ontogenetic Plasticity in Sea BassBelay AbdissaNo ratings yet
- Fish Farming ProposalDocument16 pagesFish Farming ProposalBelay Abdissa100% (4)
- Cell Science at A Glance 2681Document6 pagesCell Science at A Glance 2681Mauricio VidalNo ratings yet
- Physiological Basis of Respiratory Disease (Qutayba Hamid, Joanne Shannon MD, James Martin)Document816 pagesPhysiological Basis of Respiratory Disease (Qutayba Hamid, Joanne Shannon MD, James Martin)RLibdehNo ratings yet
- Immunology Final X 2021Document491 pagesImmunology Final X 2021nNo ratings yet
- Model Exam Round 1Document13 pagesModel Exam Round 1Gashaw ShanbelNo ratings yet
- Bls - Acls: Roli Joseph Z. Monta, RN BLS, Acls, Trauma, Hazmat CertifiedDocument72 pagesBls - Acls: Roli Joseph Z. Monta, RN BLS, Acls, Trauma, Hazmat CertifiedEkoy TheRealNo ratings yet
- Lesson - Cell Division - DifferentiationDocument8 pagesLesson - Cell Division - DifferentiationramyaNo ratings yet
- PsychologyDocument8 pagesPsychologyNikhil JosephNo ratings yet
- Gram-Negative Rods Related To The Enteric Tract The EnterobacteriaceaeDocument47 pagesGram-Negative Rods Related To The Enteric Tract The EnterobacteriaceaeShattered SoulNo ratings yet
- SyllabusDocument26 pagesSyllabuselizabethkaniparampilNo ratings yet
- Photosynthesis Light and Dark ReactionDocument36 pagesPhotosynthesis Light and Dark ReactionWHWHWHWHWWH WWGHWHGWGHWNo ratings yet
- Polycythemia Vera: DR - Karthik.S Moderator:Dr - Sumedh ShettyDocument51 pagesPolycythemia Vera: DR - Karthik.S Moderator:Dr - Sumedh ShettyDr. Apoorva KottaryNo ratings yet
- Latihan Soal Usp 2023Document10 pagesLatihan Soal Usp 2023RafelNo ratings yet
- Drug Study Isoxuprine HCL BASUEL, Aira Jane S.Document3 pagesDrug Study Isoxuprine HCL BASUEL, Aira Jane S.Aira Jane BasuelNo ratings yet
- Day 5 Cam, C3, C4Document50 pagesDay 5 Cam, C3, C4Bruh MomentNo ratings yet
- Summative Test 4th - DigestiveDocument2 pagesSummative Test 4th - DigestiveMay Joy VitorioNo ratings yet
- Cell Transport Worksheet g8Document4 pagesCell Transport Worksheet g8Ariane DionisioNo ratings yet
- Instrumentation and Quality Control in HemostasisDocument5 pagesInstrumentation and Quality Control in HemostasisGerly MaglangitNo ratings yet
- Effek Beta-Blocker Pada Sistem Kardiovaskular-Praktikum-1Document54 pagesEffek Beta-Blocker Pada Sistem Kardiovaskular-Praktikum-1Fanny MaulidaNo ratings yet
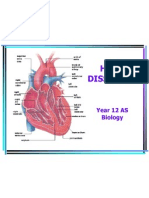
- Heart Dissection: Year 12 AS BiologyDocument9 pagesHeart Dissection: Year 12 AS BiologyBio SciencesNo ratings yet
- Actvities in General Botany Module 3Document5 pagesActvities in General Botany Module 3Ej FerrerNo ratings yet
- Y6 Math Graphs WorksheetsDocument16 pagesY6 Math Graphs WorksheetsCambridge for SuccessNo ratings yet
- Organ SystemDocument16 pagesOrgan SystemStephany RaveloNo ratings yet
- Motivation and EmotionDocument24 pagesMotivation and EmotionRebekah MathewNo ratings yet
- CT10382Document4 pagesCT10382Nguyễn HuynhNo ratings yet
- Alar Base Reduction and Alar-Columellar RelationshipDocument9 pagesAlar Base Reduction and Alar-Columellar RelationshipFabian Camelo OtorrinoNo ratings yet
- 9 - Physiology MCQ of CardiovascularDocument18 pages9 - Physiology MCQ of CardiovascularMuhammad Javed GabaNo ratings yet
- Dynamics of Insect-Plant Relationships and Its Applications in Crop ProductionDocument7 pagesDynamics of Insect-Plant Relationships and Its Applications in Crop Productionuaggallo jerryNo ratings yet
- Goodman and Gilman's Sample ChapterDocument17 pagesGoodman and Gilman's Sample Chapteradnankhan20221984No ratings yet
- Muscular System NotesDocument6 pagesMuscular System NotesZussette Corbita VingcoNo ratings yet
- Discrepant Usage of The Term Ovoviviparity' in The Herpetological LiteratureDocument9 pagesDiscrepant Usage of The Term Ovoviviparity' in The Herpetological LiteratureAndrea Dr FanisNo ratings yet