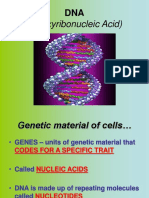
Download as pdf or txt
You might also like
- Chapter 5 ProblemsDocument3 pagesChapter 5 Problemswalt richards0% (1)
- A Level Biology A Core Practical 14 - Gel ElectrophoresisDocument3 pagesA Level Biology A Core Practical 14 - Gel ElectrophoresisAlfred Sang0% (1)
- DNA and Protein Synthesis WorksheetDocument5 pagesDNA and Protein Synthesis Worksheeteula faith miracle andam100% (2)
- 23 Sanger Sequencing SimulationDocument8 pages23 Sanger Sequencing SimulationS. SpencerNo ratings yet
- Unit 3 - Cheat Sheet Cell Enegertics PDFDocument1 pageUnit 3 - Cheat Sheet Cell Enegertics PDFJ15No ratings yet
- DNA Replication: DNA Replication Is A Biological Process That Occurs in AllDocument12 pagesDNA Replication: DNA Replication Is A Biological Process That Occurs in AllAnonymous HChxHbANo ratings yet
- Central DogmaDocument4 pagesCentral DogmaRodriguez MiaNo ratings yet
- Unit 5 Notes Biology AqaDocument94 pagesUnit 5 Notes Biology Aqa李超然No ratings yet
- Dna Sequencing (Sanger's Method)Document12 pagesDna Sequencing (Sanger's Method)Ajay KumarNo ratings yet
- HM 11Document51 pagesHM 11body fayezNo ratings yet
- Cell Structure and FunctionDocument41 pagesCell Structure and FunctionAhkyluzLaniaz100% (1)
- NNNDocument27 pagesNNNAnwar AliNo ratings yet
- Replication, Transcription, Translation, Mutation HWDocument20 pagesReplication, Transcription, Translation, Mutation HWtahamidNo ratings yet
- 07 Nucleic Acids Test PDFDocument8 pages07 Nucleic Acids Test PDFUkjun JungNo ratings yet
- Prepared By: Verna Jean M. Magdayao 3/Bsbiology/ADocument56 pagesPrepared By: Verna Jean M. Magdayao 3/Bsbiology/AKathleya PeñaNo ratings yet
- S.Y.Bsc Semester Iii Botany Paper IiDocument53 pagesS.Y.Bsc Semester Iii Botany Paper IiĐỗ Quang BìnhNo ratings yet
- Biology Key TermsDocument3 pagesBiology Key TermsIshra ChaudhryNo ratings yet
- Sequecing - Khan AcademyDocument16 pagesSequecing - Khan AcademyMaryem SafdarNo ratings yet
- Biology A Level OCR 2022Document6 pagesBiology A Level OCR 2022shNo ratings yet
- Restriction Enzyme DigestionDocument5 pagesRestriction Enzyme DigestionAqsa ImtiazNo ratings yet
- Experiment No.-6 Agarose Gel ElectrophoresisDocument10 pagesExperiment No.-6 Agarose Gel ElectrophoresisAditya JhaNo ratings yet
- DNA Analysis Sanger Sequencing: SeminarDocument51 pagesDNA Analysis Sanger Sequencing: SeminarSV. Trương Nguyễn Hoàng LongNo ratings yet
- Restriction Enzyme Digestion of DNADocument2 pagesRestriction Enzyme Digestion of DNAMustansir BhoriNo ratings yet
- Inheritance Chap 17Document23 pagesInheritance Chap 17Rayan KhanNo ratings yet
- Deoxyribonucleic AcidDocument36 pagesDeoxyribonucleic AcidAlex LlanoNo ratings yet
- Dna and Rna StructureDocument63 pagesDna and Rna StructureKhaled DabourNo ratings yet
- Cell Cycle Cell Division PDFDocument13 pagesCell Cycle Cell Division PDFmuzamil shabir100% (1)
- Unit 4 Revision Notes SNAB Topics 5 & 6 A2Document38 pagesUnit 4 Revision Notes SNAB Topics 5 & 6 A2Olivia Panterka Vainilla100% (2)
- Cell Biology Practice 2Document15 pagesCell Biology Practice 2NgMinhHaiNo ratings yet
- DNA Replication in ProkaryotesDocument24 pagesDNA Replication in ProkaryotesAlbert Jade Pontimayor Legaria100% (1)
- Classification StudysheetDocument36 pagesClassification Studysheetwafa elias100% (1)
- Lecture 3: DNA RecombinationDocument22 pagesLecture 3: DNA RecombinationBiotechnology IIUM Kuantan100% (2)
- Homework 7Document62 pagesHomework 7body fayezNo ratings yet
- Prokaryoticneukaryoticgenome 160514180458 PDFDocument7 pagesProkaryoticneukaryoticgenome 160514180458 PDFrahul ssgiNo ratings yet
- Assignment of Molecular BiologyDocument4 pagesAssignment of Molecular BiologyNasir Hussain FarazNo ratings yet
- Practice Test 2Document5 pagesPractice Test 2crazybobblaskeyNo ratings yet
- Biology DefinitionsDocument6 pagesBiology DefinitionsSixtine MorinNo ratings yet
- Cloning VectorsDocument8 pagesCloning Vectorssanchit100% (3)
- Variation and GeneticsDocument11 pagesVariation and GeneticsTasawar Hussain DayoNo ratings yet
- DNA Sequencing LectureDocument29 pagesDNA Sequencing LectureBalew GetaNo ratings yet
- Molecular Biology BIOL312Document273 pagesMolecular Biology BIOL312Paul GarciaNo ratings yet
- RNA Protein Synthesis Gizmo COMPLETEDDocument7 pagesRNA Protein Synthesis Gizmo COMPLETEDcokcreNo ratings yet
- Extraction of Genomic DNA: G.Umamaheswaran PH.D Scholar JipmerDocument24 pagesExtraction of Genomic DNA: G.Umamaheswaran PH.D Scholar Jipmerpharmaguy111No ratings yet
- DNA ExtractionDocument5 pagesDNA ExtractionDana Porter100% (1)
- Unit 3 EvolutionDocument51 pagesUnit 3 EvolutionFatma Taha MohamedNo ratings yet
- Nucleic AcidsDocument36 pagesNucleic AcidsAman UllahNo ratings yet
- Avatics 2017 03-02-15!34!57protein Synthesis Worksheet PracticeDocument2 pagesAvatics 2017 03-02-15!34!57protein Synthesis Worksheet PracticeMiguel BernalNo ratings yet
- Cell Structure: Question Paper 1Document12 pagesCell Structure: Question Paper 1rkblsistemNo ratings yet
- Regulation of Histidine and Hut OperonsDocument11 pagesRegulation of Histidine and Hut Operonsaditi_joshee419No ratings yet
- Heredity and VariationDocument79 pagesHeredity and VariationNathiya100% (1)
- DNA Repair: Chapter FourDocument7 pagesDNA Repair: Chapter FourMatin Ahmad KhanNo ratings yet
- Xu Zhaoyang Department of Cell Biology Basic Medical College Zhengzhou UniversityDocument35 pagesXu Zhaoyang Department of Cell Biology Basic Medical College Zhengzhou Universityapi-19916399100% (1)
- Epithelial TissueDocument87 pagesEpithelial Tissueabubeker mohammed100% (1)
- Linkage & Chromosome MappingDocument35 pagesLinkage & Chromosome MappingTouhidulIslamNo ratings yet
- DNA SequencingDocument21 pagesDNA SequencingGanesh V GaonkarNo ratings yet
- 7th Cytogenetics Lecture 20121 1Document37 pages7th Cytogenetics Lecture 20121 1RC SILVESTRENo ratings yet
- The Central Dogma of Molecular BiologyDocument5 pagesThe Central Dogma of Molecular BiologyMartires, Vea Janela P.100% (1)
- Biology Chapter 21 Sylvia Mader ModifiedDocument29 pagesBiology Chapter 21 Sylvia Mader ModifiedabdallahNo ratings yet
- A Local EcosystemDocument26 pagesA Local EcosystemsandnifflerNo ratings yet
- Recombinant Dna Technology Project Based Learning Topic-: Sanger SequencingDocument6 pagesRecombinant Dna Technology Project Based Learning Topic-: Sanger Sequencingvidushi srivastavaNo ratings yet
- 2016 Immunity2009Document42 pages2016 Immunity2009api-277471896No ratings yet
- Nervous System 2016Document31 pagesNervous System 2016api-277471896No ratings yet
- Generation of Diversity in BacteriaDocument12 pagesGeneration of Diversity in Bacteriaapi-277471896No ratings yet
- Prokaryotic Gene Regulation NotesDocument16 pagesProkaryotic Gene Regulation Notesapi-277471896No ratings yet
- Genetoprotein FogliaDocument45 pagesGenetoprotein Fogliaapi-277471896100% (1)
- WaterpotentialDocument18 pagesWaterpotentialapi-277471896No ratings yet
- Eukaryotic Gene Regulation NotesDocument16 pagesEukaryotic Gene Regulation Notesapi-277471896No ratings yet
- Regulation of Cell Division: AP BiologyDocument26 pagesRegulation of Cell Division: AP Biologyapi-277471896No ratings yet
- Dna Structure ReplicationDocument34 pagesDna Structure Replicationapi-277471896No ratings yet
- Metabolism ReviewDocument20 pagesMetabolism Reviewapi-277471896No ratings yet
- Cell Intro Notes - Folglia ModifiedDocument14 pagesCell Intro Notes - Folglia Modifiedapi-277471896No ratings yet
- 02 CH 05 Organelles 12007Document20 pages02 CH 05 Organelles 12007api-277471896No ratings yet
- Cell Communication 1Document10 pagesCell Communication 1api-277471896No ratings yet
- Intermediate Inheritance PatternsDocument29 pagesIntermediate Inheritance Patternsapi-277471896No ratings yet
- Intermediate Inheritance PatternsDocument29 pagesIntermediate Inheritance Patternsapi-277471896No ratings yet
- Carbon Powerpoint-2Document25 pagesCarbon Powerpoint-2api-277471896No ratings yet
- Chi-Squared Analysis PDF WebsiteDocument15 pagesChi-Squared Analysis PDF Websiteapi-277471896No ratings yet
- Design An Experiment: If ThenDocument2 pagesDesign An Experiment: If Thenapi-277471896No ratings yet
- Blood TypesDocument12 pagesBlood Typesapi-277471896No ratings yet
- Human ImpactDocument12 pagesHuman Impactapi-277471896No ratings yet
- Animal BehaviorDocument13 pagesAnimal Behaviorapi-277471896No ratings yet
- Molecular Genetics Review Notes PDFDocument29 pagesMolecular Genetics Review Notes PDFapi-277471896No ratings yet
- Protien SynthesisDocument20 pagesProtien Synthesisapi-277471896No ratings yet
- Ap Biology Chapter 22 DarwinDocument22 pagesAp Biology Chapter 22 Darwinapi-277471896No ratings yet
- Osmosis NotesDocument8 pagesOsmosis Notesapi-277471896No ratings yet
- Electrical Discharge MachiningDocument11 pagesElectrical Discharge MachiningRizwan MrnNo ratings yet
- Sales Management Course Panda Sachdev 2259Document1,408 pagesSales Management Course Panda Sachdev 2259raachyeta_sharmaNo ratings yet
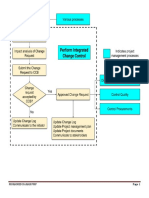
- Perform Integrated Change Control (Step-By-Step)Document4 pagesPerform Integrated Change Control (Step-By-Step)Justin M. FowlkesNo ratings yet
- Contract Management LectureDocument84 pagesContract Management LectureYasin ShahNo ratings yet
- MODULE 2: Assessment #2 SCI 1: Teaching Science in The Elementary Grades-Biology and ChemistryDocument6 pagesMODULE 2: Assessment #2 SCI 1: Teaching Science in The Elementary Grades-Biology and Chemistryshine brightNo ratings yet
- Loibl Bacher BGP Flowspec Interop 012017Document64 pagesLoibl Bacher BGP Flowspec Interop 012017Antonio VieiraNo ratings yet
- Professional Practice-Ii Law of Land: Prepared By: - Bhoomit Kansara Yash Gosai Sahil Vora Part SisodiyaDocument19 pagesProfessional Practice-Ii Law of Land: Prepared By: - Bhoomit Kansara Yash Gosai Sahil Vora Part SisodiyaMayur KotechaNo ratings yet
- CASIODocument9 pagesCASIOThomas GeorgeNo ratings yet
- Inverted Tubes 2Document3 pagesInverted Tubes 2tihomihoNo ratings yet
- Steyn City BrochureDocument36 pagesSteyn City BrochureCorey AcevedoNo ratings yet
- Cot DLP Oral Com q1-2019Document1 pageCot DLP Oral Com q1-2019monette refuerzo0% (1)
- A System View To Urban Planning: An IntroductionDocument34 pagesA System View To Urban Planning: An IntroductionAzad RazaNo ratings yet
- BF422 DatasheetDocument4 pagesBF422 DatasheetCarlos GuilarteNo ratings yet
- Semester IIDocument11 pagesSemester IIAnnantNo ratings yet
- Intuitive Surgical JP Morgan PresentationDocument25 pagesIntuitive Surgical JP Morgan PresentationmedtechyNo ratings yet
- Introduction To ProgrammingDocument104 pagesIntroduction To ProgrammingMiliyon TilahunNo ratings yet
- Following Are The Operations On Segment Creating The SegmentDocument2 pagesFollowing Are The Operations On Segment Creating The Segmentabhishek125No ratings yet
- Sample Detailed Lesson PlanDocument6 pagesSample Detailed Lesson PlanKarlKarl100% (1)
- Nwda Je Ce 2019 EnglishDocument24 pagesNwda Je Ce 2019 EnglishHarsha Vardhan KoppulaNo ratings yet
- Posse. Analecta Vaticana. 1878.Document240 pagesPosse. Analecta Vaticana. 1878.Patrologia Latina, Graeca et OrientalisNo ratings yet
- Kill Team Campaign Cheat SheetDocument3 pagesKill Team Campaign Cheat SheetangelofmenothNo ratings yet
- MCQ On OBDocument21 pagesMCQ On OBRishikesh PatilNo ratings yet
- Material Safety Data SheetDocument4 pagesMaterial Safety Data SheetIvani SoaresNo ratings yet
- Congress Program - Updated 09.05.2018Document4 pagesCongress Program - Updated 09.05.2018Booker DewittNo ratings yet
- HMBA Design DossierDocument28 pagesHMBA Design DossierDaniel BarrenecheaNo ratings yet
- Allison 4th Generation ControlsDocument22 pagesAllison 4th Generation Controlsmnvijaybabu100% (1)
- Daily Task of Disbursing OfficerDocument2 pagesDaily Task of Disbursing OfficerEijneb OptionalNo ratings yet
- 10 CarbohydratesDocument6 pages10 CarbohydratesBryanNo ratings yet
- Armstrong Flute Brochure PDFDocument8 pagesArmstrong Flute Brochure PDFpoptot100% (1)
- Topsolid 2014 What'S New in Topsolid'Design V6.15Document12 pagesTopsolid 2014 What'S New in Topsolid'Design V6.15Nina GregorioNo ratings yet