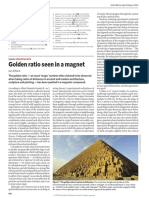
Download as pdf or txt
You might also like
- Workflow - Technical Design TemplateDocument10 pagesWorkflow - Technical Design TemplateKrishna MurthyNo ratings yet
- Kattnig and Hore - The Sensitivity of A Radical Pair Compass Magnetoreceptor Can Be Significantly Amplified by Radical ScavengersDocument24 pagesKattnig and Hore - The Sensitivity of A Radical Pair Compass Magnetoreceptor Can Be Significantly Amplified by Radical Scavengersalt6No ratings yet
- The Quantum Needle of The Avian Magnetic Compass: SignificanceDocument6 pagesThe Quantum Needle of The Avian Magnetic Compass: SignificanceBenancio DorantessNo ratings yet
- Stoneham Et Al. - 2012 - A New Type of Radical-Pair-Based Model For MagnetoreceptionDocument8 pagesStoneham Et Al. - 2012 - A New Type of Radical-Pair-Based Model For Magnetoreceptionalt6No ratings yet
- Geomagnetic CryptochromeDocument10 pagesGeomagnetic CryptochromeordoNo ratings yet
- 10.1038@s41586 020 2051 0Document16 pages10.1038@s41586 020 2051 0gabrielpoulsonNo ratings yet
- Molecular Dynamics Simulations Disclose Early Stages of The Photo-Activation of Cryptochrome 4Document19 pagesMolecular Dynamics Simulations Disclose Early Stages of The Photo-Activation of Cryptochrome 4GwainNo ratings yet
- 2013 - PRA. Spontaneous Emission in The Presence of A Spherical Plasmonic MetamaterialDocument7 pages2013 - PRA. Spontaneous Emission in The Presence of A Spherical Plasmonic MetamaterialDiego OliverNo ratings yet
- 1807.01822 - Correlações de Spin Termicamente Robustas Entre Dois Átomos de 85Rb em Um Microtrap ÓpticoDocument18 pages1807.01822 - Correlações de Spin Termicamente Robustas Entre Dois Átomos de 85Rb em Um Microtrap ÓpticomarcelokbeloNo ratings yet
- Nihms57127 Magnetosensitivity in DrosophilaDocument14 pagesNihms57127 Magnetosensitivity in Drosophila21lbph02No ratings yet
- Direct Assignment of Molecular Vibrations Via Normal Mode Analysis of The Neutron Dynamic Pair Distribution Function TechniqueDocument24 pagesDirect Assignment of Molecular Vibrations Via Normal Mode Analysis of The Neutron Dynamic Pair Distribution Function TechniqueRabbia AminNo ratings yet
- Thorsten Ritz, Salih Adem, Klaus Schulten - A Model For Photoreceptor-Based Magnetoreception in Birds PDFDocument12 pagesThorsten Ritz, Salih Adem, Klaus Schulten - A Model For Photoreceptor-Based Magnetoreception in Birds PDFwise monkeyNo ratings yet
- Spin Selective Transport Through Helical Molecular Systems: PACS Numbers: 73.22.-f 73.63.-b 72.25.-b 87.14.gk 87.15.PcDocument4 pagesSpin Selective Transport Through Helical Molecular Systems: PACS Numbers: 73.22.-f 73.63.-b 72.25.-b 87.14.gk 87.15.PcymonsalveNo ratings yet
- Bem 20485Document9 pagesBem 20485Malinus TorquatusNo ratings yet
- Theory of High Energy Optical Conductivity and The Role of Oxygens in ManganitesDocument8 pagesTheory of High Energy Optical Conductivity and The Role of Oxygens in ManganitesAslamic le ChauveNo ratings yet
- Superconductivity and Strong Interactions in A Tunable Moir e Quasiperiodic CrystalDocument25 pagesSuperconductivity and Strong Interactions in A Tunable Moir e Quasiperiodic Crystalalex moreno sanchezNo ratings yet
- Characterizing The Effect of Static Magnetic Fields On MicroorganismsDocument9 pagesCharacterizing The Effect of Static Magnetic Fields On MicroorganismsSantosh PandeyNo ratings yet
- Energy-Density Functional Plus Quasiparticle-Phonon Model Theory As A Powerful Tool For Nuclear Structure and AstrophysicsDocument19 pagesEnergy-Density Functional Plus Quasiparticle-Phonon Model Theory As A Powerful Tool For Nuclear Structure and AstrophysicsNishantShuklaNo ratings yet
- Berrada 2023 Laser Phys. Lett. 20 085201Document7 pagesBerrada 2023 Laser Phys. Lett. 20 085201Hichem El EuchNo ratings yet
- PhysRevB 100 024429Document10 pagesPhysRevB 100 024429jitenderthakurNo ratings yet
- Lifson 1957Document9 pagesLifson 1957brouuorbNo ratings yet
- Ampc 2023092016315868Document10 pagesAmpc 2023092016315868Putu IndraNo ratings yet
- Austin 2015Document17 pagesAustin 2015Premkumar RNo ratings yet
- Phy Revb PDFDocument11 pagesPhy Revb PDFVignesh RamakrishnanNo ratings yet
- Prentiss - Dissociation of Ligand-Receptor Complexes Using Magnetic TweezersDocument6 pagesPrentiss - Dissociation of Ligand-Receptor Complexes Using Magnetic TweezersChris TranNo ratings yet
- Articles: Detailed Mechanism For Trans-Cis Photoisomerization of Butadiene Following A Femtosecond-Scale Laser PulseDocument8 pagesArticles: Detailed Mechanism For Trans-Cis Photoisomerization of Butadiene Following A Femtosecond-Scale Laser PulseSigid Boedy A SGNo ratings yet
- Anisotropic Destruction of The Fermi Surface in Inhomogeneous Holographic LatticesDocument25 pagesAnisotropic Destruction of The Fermi Surface in Inhomogeneous Holographic LatticesAndrey BagrovNo ratings yet
- Polar PNA Exp PDFDocument13 pagesPolar PNA Exp PDFIgor GornéNo ratings yet
- ElectrochemistryDocument3 pagesElectrochemistryakibNo ratings yet
- Kramer 1995Document14 pagesKramer 1995elkadaouisoukainaNo ratings yet
- Paper Revision ModelDocument15 pagesPaper Revision ModelJason Jerry Atoche MedranoNo ratings yet
- Toward An Accurate Strongly-Coupled Many-Body Theory Within The Equation of Motion FrameworkDocument24 pagesToward An Accurate Strongly-Coupled Many-Body Theory Within The Equation of Motion FrameworkGaston GBNo ratings yet
- Symbiotic Magnetic Sensing: Raising Evidence and Beyond: ReviewDocument8 pagesSymbiotic Magnetic Sensing: Raising Evidence and Beyond: Reviewarie.zychlinskiNo ratings yet
- Meeting Reviews: Quantum Mechanics and Chemistry: The Relevance of Nonlocality and Entangle-Ment For MoleculesDocument2 pagesMeeting Reviews: Quantum Mechanics and Chemistry: The Relevance of Nonlocality and Entangle-Ment For MoleculesIan FreemanNo ratings yet
- Magnetic-Field-Induced Delocalization in Hybrid Electron-Nuclear Spin EnsemblesDocument14 pagesMagnetic-Field-Induced Delocalization in Hybrid Electron-Nuclear Spin Ensemblescharushila patelNo ratings yet
- Formation of Buried Domain Walls in The Ultrafast Transition of SmteDocument6 pagesFormation of Buried Domain Walls in The Ultrafast Transition of SmteErick ChangNo ratings yet
- Magnetic Sensitivity of Cryptochrome 4 From A Migratory SongbirdDocument9 pagesMagnetic Sensitivity of Cryptochrome 4 From A Migratory SongbirdBhanu SinghNo ratings yet
- Allain Comm Nat MatDocument2 pagesAllain Comm Nat MatleNo ratings yet
- Pac Con 08 10 18 - 230305 - 180556Document26 pagesPac Con 08 10 18 - 230305 - 180556Mohamed TarekNo ratings yet
- Charge Transfer Conjugated Polymers Science 2009Document3 pagesCharge Transfer Conjugated Polymers Science 2009PengHanNo ratings yet
- 2008 JPhys CMSpaldinDocument16 pages2008 JPhys CMSpaldinXavier CazorNo ratings yet
- Electronic G Tensors in U Complexes-A Computational Study: DOI: 10.1002/chem.201701058Document11 pagesElectronic G Tensors in U Complexes-A Computational Study: DOI: 10.1002/chem.201701058Austin LloydNo ratings yet
- Affleck2010 PDFDocument2 pagesAffleck2010 PDFAnsor Prima YudaNo ratings yet
- Srep 07892Document7 pagesSrep 07892ResalatFaruqueNuhNo ratings yet
- Transmission by Solitary Waves in Biological Systems: LeninskyDocument11 pagesTransmission by Solitary Waves in Biological Systems: LeninskyLejirbag CevardazNo ratings yet
- PhysRevResearch 2 023131Document15 pagesPhysRevResearch 2 023131enrique hernandezNo ratings yet
- 1 6 Williamson1994Document16 pages1 6 Williamson1994Hien NguyenNo ratings yet
- Nikolaev - 2016 - J. - Phys. - Conf. - Ser. - 774 - 012172Document7 pagesNikolaev - 2016 - J. - Phys. - Conf. - Ser. - 774 - 012172Li ChNo ratings yet
- Microscopic Artificial Swimmers - Rémi Dreyfus Et AlDocument5 pagesMicroscopic Artificial Swimmers - Rémi Dreyfus Et Alalfredc20No ratings yet
- Science at Ultra-Brieftime Periods From The Attosecond To The PetawattDocument8 pagesScience at Ultra-Brieftime Periods From The Attosecond To The PetawattPocxaNo ratings yet
- Recent Progress and Prospects in Plasmon-Mediated Chemical ReactionDocument15 pagesRecent Progress and Prospects in Plasmon-Mediated Chemical ReactionNurdan AşarNo ratings yet
- 1304 5379v1 PDFDocument5 pages1304 5379v1 PDFpippoaNo ratings yet
- 1989.04.01 LIU PRB MagnetismDocument11 pages1989.04.01 LIU PRB MagnetismAlejandra AwimbaweNo ratings yet
- Reacción en UltrafrioDocument6 pagesReacción en UltrafrioDaniel Alejandro Rojas ToroNo ratings yet
- 13 Forrester Switching Dynamics of Doped CoFeB Trilayers PhysRevB.87.174416Document17 pages13 Forrester Switching Dynamics of Doped CoFeB Trilayers PhysRevB.87.174416mgrabows18No ratings yet
- Biskup Et Al-2009-Angewandte Chemie (International Ed. in English)Document4 pagesBiskup Et Al-2009-Angewandte Chemie (International Ed. in English)Marco MijaresNo ratings yet
- The Future of Quantum Biology: Headline ReviewDocument14 pagesThe Future of Quantum Biology: Headline Reviewcristian flore arreolaNo ratings yet
- Sunday Afternoon, October 27, 2013: TechnologyDocument283 pagesSunday Afternoon, October 27, 2013: TechnologyNatasha MyersNo ratings yet
- Strongly Interacting Polaritons in Coupled Arrays of CavitiesDocument7 pagesStrongly Interacting Polaritons in Coupled Arrays of CavitiesJulien KuntzNo ratings yet
- Magnetic Reconnection: A Modern Synthesis of Theory, Experiment, and ObservationsFrom EverandMagnetic Reconnection: A Modern Synthesis of Theory, Experiment, and ObservationsNo ratings yet
- On the Revolutions of the Internal Spheres: A New Theory of Matter and the Transmission of Light, Second EditionFrom EverandOn the Revolutions of the Internal Spheres: A New Theory of Matter and the Transmission of Light, Second EditionNo ratings yet
- Noise-Assisted Energy Transfer in Quantum Networks and Light-Harvesting ComplexesDocument17 pagesNoise-Assisted Energy Transfer in Quantum Networks and Light-Harvesting Complexesgaminster6935No ratings yet
- Open Quantum System Approach To The Modeling of Spin Recombination ReactionsDocument20 pagesOpen Quantum System Approach To The Modeling of Spin Recombination Reactionsgaminster6935No ratings yet
- Probing Biological Light-Harvesting Phenomena by Optical CavitiesDocument10 pagesProbing Biological Light-Harvesting Phenomena by Optical Cavitiesgaminster6935No ratings yet
- Quantum Transport in Quantum Networks and Photosynthetic Complexes at The Steady StateDocument9 pagesQuantum Transport in Quantum Networks and Photosynthetic Complexes at The Steady Stategaminster6935No ratings yet
- Persistent Dynamic Entanglement From Classical Motion: How Bio-Molecular Machines Can Generate Nontrivial Quantum StatesDocument22 pagesPersistent Dynamic Entanglement From Classical Motion: How Bio-Molecular Machines Can Generate Nontrivial Quantum Statesgaminster6935No ratings yet
- Dissipation and Decoherence in Quantum Systems: F. Marquardt and A. Püttmann February 4, 2008Document21 pagesDissipation and Decoherence in Quantum Systems: F. Marquardt and A. Püttmann February 4, 2008gaminster6935No ratings yet
- DT 1203.5072v1Document12 pagesDT 1203.5072v1gaminster6935No ratings yet
- Quantum Coherence in Photosynthesis For Efficient Solar-Energy ConversionDocument7 pagesQuantum Coherence in Photosynthesis For Efficient Solar-Energy Conversiongaminster6935No ratings yet
- MIT DephasinNoiseDocument19 pagesMIT DephasinNoisegaminster6935No ratings yet
- A Practical Method For Calculating Largest Lyapunov Exponents From Small Data SetsDocument18 pagesA Practical Method For Calculating Largest Lyapunov Exponents From Small Data Setsgaminster6935No ratings yet
- Dephasing-Assisted Transport: Quantum Networks and BiomoleculesDocument15 pagesDephasing-Assisted Transport: Quantum Networks and Biomoleculesgaminster6935No ratings yet
- Dynamic Entanglement in Oscillating Molecules and Potential Biological ImplicationsDocument10 pagesDynamic Entanglement in Oscillating Molecules and Potential Biological Implicationsgaminster6935No ratings yet
- Decoherence in Strongly Coupled Quantum Oscillators: M.A. de Ponte, M.C. de Oliveira, M.H.Y. MoussaDocument35 pagesDecoherence in Strongly Coupled Quantum Oscillators: M.A. de Ponte, M.C. de Oliveira, M.H.Y. Moussagaminster6935No ratings yet
- Administrative Mishap (Supergirl - Worm) - SpaceBattlesDocument114 pagesAdministrative Mishap (Supergirl - Worm) - SpaceBattlesgaminster6935No ratings yet
- Steady-State Entanglement in Open and Noisy Quantum Systems: 10.1103/physreva.74.052304Document5 pagesSteady-State Entanglement in Open and Noisy Quantum Systems: 10.1103/physreva.74.052304gaminster6935No ratings yet
- Inspired Voyage (ST - Voyager - SI) - Page 5 - SpaceBattlesDocument100 pagesInspired Voyage (ST - Voyager - SI) - Page 5 - SpaceBattlesgaminster6935No ratings yet
- Inspired Voyage (ST - Voyager - SI) - SpaceBattlesDocument96 pagesInspired Voyage (ST - Voyager - SI) - SpaceBattlesgaminster6935100% (1)
- Administrative Mishap (Supergirl - Worm) - Page 2 - SpaceBattlesDocument127 pagesAdministrative Mishap (Supergirl - Worm) - Page 2 - SpaceBattlesgaminster6935No ratings yet
- Inspired Voyage (ST - Voyager - SI) - Page 3 - SpaceBattlesDocument118 pagesInspired Voyage (ST - Voyager - SI) - Page 3 - SpaceBattlesgaminster6935No ratings yet
- Inspired Voyage (ST - Voyager - SI) - Page 2 - SpaceBattlesDocument88 pagesInspired Voyage (ST - Voyager - SI) - Page 2 - SpaceBattlesgaminster6935No ratings yet
- Inspired Voyage (ST - Voyager - SI) - Page 4 - SpaceBattlesDocument103 pagesInspired Voyage (ST - Voyager - SI) - Page 4 - SpaceBattlesgaminster6935No ratings yet
- Administrative Mishap (Supergirl - Worm) - Page 7 - SpaceBattlesDocument162 pagesAdministrative Mishap (Supergirl - Worm) - Page 7 - SpaceBattlesgaminster6935No ratings yet
- Administrative Mishap (Supergirl - Worm) - Page 5 - SpaceBattlesDocument153 pagesAdministrative Mishap (Supergirl - Worm) - Page 5 - SpaceBattlesgaminster6935No ratings yet
- Lj14115e PDFDocument9 pagesLj14115e PDFgaminster6935No ratings yet
- Administrative Mishap (Supergirl - Worm) - Page 3 - SpaceBattlesDocument119 pagesAdministrative Mishap (Supergirl - Worm) - Page 3 - SpaceBattlesgaminster6935No ratings yet
- Administrative Mishap (Supergirl - Worm) - Page 6 - SpaceBattlesDocument148 pagesAdministrative Mishap (Supergirl - Worm) - Page 6 - SpaceBattlesgaminster6935No ratings yet
- Administrative Mishap (Supergirl - Worm) - Page 4 - SpaceBattlesDocument144 pagesAdministrative Mishap (Supergirl - Worm) - Page 4 - SpaceBattlesgaminster6935No ratings yet
- Inspired Voyage (ST - Voyager - SI) - Page 6 - SpaceBattlesDocument66 pagesInspired Voyage (ST - Voyager - SI) - Page 6 - SpaceBattlesgaminster6935No ratings yet
- Carbocation - Wikipedia, The Free EncyclopediaDocument5 pagesCarbocation - Wikipedia, The Free EncyclopediaBenni WewokNo ratings yet
- GRE Verbal Reasoning Questions and AnswersDocument24 pagesGRE Verbal Reasoning Questions and AnswersTamunotonye Minaye-IlomboNo ratings yet
- Code Addicts - The Python Starter Kit - An In-Depth and Practical Course For Beginners To Python Programming. Including Detailed Step-By-step Guides and Practical Demonstrations. (2017)Document136 pagesCode Addicts - The Python Starter Kit - An In-Depth and Practical Course For Beginners To Python Programming. Including Detailed Step-By-step Guides and Practical Demonstrations. (2017)Roland Rawlins Igabor100% (2)
- Mathematics 7 SECOND QUARTEER (S.Y. 2020-2021) Weekly-Home Learning PlanDocument3 pagesMathematics 7 SECOND QUARTEER (S.Y. 2020-2021) Weekly-Home Learning PlanlyspinNo ratings yet
- Dynamic Positioning Revalidation: Online Application GuideDocument23 pagesDynamic Positioning Revalidation: Online Application GuideCandisftNo ratings yet
- SBSTC & RedBus General Terms and ConditionsDocument11 pagesSBSTC & RedBus General Terms and ConditionsAjoy RoyNo ratings yet
- Waterwheels Information PaperDocument4 pagesWaterwheels Information PaperCat MaNo ratings yet
- Eco 270Document9 pagesEco 270Tarun Shankar ChoudharyNo ratings yet
- Mobile Phone: Service ManualDocument128 pagesMobile Phone: Service ManualsamuelNo ratings yet
- Cal Am Manufacturing Digital CatalogDocument42 pagesCal Am Manufacturing Digital CatalogeasyxcessNo ratings yet
- Administration and ManagementDocument28 pagesAdministration and ManagementMonikaNo ratings yet
- SBQ On Globalization in SingaporeDocument4 pagesSBQ On Globalization in SingaporeMegan NgNo ratings yet
- Autonomous Systems: Inertial SystemsDocument50 pagesAutonomous Systems: Inertial SystemsSara Nieves Matheu GarciaNo ratings yet
- Sepharial - The Manual of Astrology - Book III The Measure of TimeDocument42 pagesSepharial - The Manual of Astrology - Book III The Measure of TimeJade Willowbean100% (1)
- Clean Technology in The Production of Epichlorohydrin: Jowi W. Bijsterbosch, A. Das and F.P.J.M. KerkhofDocument4 pagesClean Technology in The Production of Epichlorohydrin: Jowi W. Bijsterbosch, A. Das and F.P.J.M. KerkhofProton HutabaratNo ratings yet
- Dental TEST-BANKDocument11 pagesDental TEST-BANKMC EllisNo ratings yet
- Literature Review Scope ExampleDocument5 pagesLiterature Review Scope Exampleea20cqyt100% (1)
- Regina D Perry CV 2015Document8 pagesRegina D Perry CV 2015api-284587833No ratings yet
- A The TempestDocument10 pagesA The TempestBouchra BouchraNo ratings yet
- 10x8 Shed PlanDocument11 pages10x8 Shed Planaweed100% (1)
- Lesson 2 - Duties of A Painting (Coatings) InspectorDocument55 pagesLesson 2 - Duties of A Painting (Coatings) InspectorFarhad AsadulovNo ratings yet
- ConTEXt - Installing Expert Fonts - Minion ProDocument17 pagesConTEXt - Installing Expert Fonts - Minion ProGuy StylesNo ratings yet
- The Recruitment Process: CoverDocument19 pagesThe Recruitment Process: Coveradhitiya pNo ratings yet
- Mathey Wireline UnitsDocument1 pageMathey Wireline UnitsTri CahyadiNo ratings yet
- Paper Name and Code - Sem - I ND2023Document6 pagesPaper Name and Code - Sem - I ND2023Sneha KhapraNo ratings yet
- High Lift Hydraulic Transmission Jacks "AF25082" "AF50080" "AF100080"Document6 pagesHigh Lift Hydraulic Transmission Jacks "AF25082" "AF50080" "AF100080"Alain DefoeNo ratings yet
- The Affluent Society and A Sense of EmptinessDocument31 pagesThe Affluent Society and A Sense of EmptinessMehwish MalikNo ratings yet
- Parametric Kerf Bending - ArticleDocument25 pagesParametric Kerf Bending - ArticleMIS-TRYNo ratings yet
- Catálogo Del Transformador WEGDocument12 pagesCatálogo Del Transformador WEGcarlos alfredo garcia bellidoNo ratings yet