Download as pdf or txt
You might also like
- Georg Henrik Von Wright Norm and Action 0Document201 pagesGeorg Henrik Von Wright Norm and Action 0Garai Zsolt100% (1)
- P1 Question Paper January 21Document32 pagesP1 Question Paper January 21Shere AhmedNo ratings yet
- Biol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationDocument49 pagesBiol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationDocument49 pagesBiol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationHui Ka HoNo ratings yet
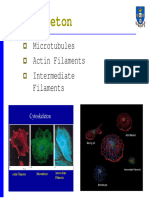
- The CytoskeletonDocument31 pagesThe CytoskeletonSanthoshsv 143No ratings yet
- Lectures 8-10 NotesDocument15 pagesLectures 8-10 Notesapi-3721438No ratings yet
- 2 Cytoskeleton & Cell MotilityDocument47 pages2 Cytoskeleton & Cell MotilityChristineNo ratings yet
- C & M Bio CytoskeletonDocument40 pagesC & M Bio CytoskeletonA PNo ratings yet
- L 4 Cell Organization IIDocument35 pagesL 4 Cell Organization IIsNo ratings yet
- Microtubule Oranizing CenterDocument5 pagesMicrotubule Oranizing CenterSasha GutierrezNo ratings yet
- CytoskeletonDocument65 pagesCytoskeletonSapana SubediNo ratings yet
- Cytoskeleton Motor Proteins and ECM - SCDocument77 pagesCytoskeleton Motor Proteins and ECM - SCAlice RizzoNo ratings yet
- MBB231 Chapter 9 Cytoskeleton Introduction 1197Document28 pagesMBB231 Chapter 9 Cytoskeleton Introduction 1197nathanNo ratings yet
- 1 7 CytoskeletonDocument31 pages1 7 CytoskeletonAditya AgrawalNo ratings yet
- Cytoskeleton DADocument35 pagesCytoskeleton DAAnonymous NVBCqKWHNo ratings yet
- DJ (Microtubules, Microfilaments and Intermediate Filaments) - DJGFDocument43 pagesDJ (Microtubules, Microfilaments and Intermediate Filaments) - DJGFNick_989893No ratings yet
- BM 1 1 CytoskeletonDocument24 pagesBM 1 1 CytoskeletonSanthoshi Sadhanaa SankarNo ratings yet
- The Cytoskeleton #1: Cytoskeletal FilamentsDocument42 pagesThe Cytoskeleton #1: Cytoskeletal FilamentsKimberly ThomasNo ratings yet
- Cytoplasm and InclusionDocument21 pagesCytoplasm and Inclusionelphas walelaNo ratings yet
- Mekanisme Kerja Organla SelDocument51 pagesMekanisme Kerja Organla SelAuliya SyismaNo ratings yet
- Nov1 Lecture For BiologyDocument30 pagesNov1 Lecture For BiologytimakuNo ratings yet
- 01 02 Cytoskeleton PDFDocument12 pages01 02 Cytoskeleton PDFsigninNo ratings yet
- Enumerating The Structure of Major Protein, Describe The Eukaryotic CytoskeletonDocument10 pagesEnumerating The Structure of Major Protein, Describe The Eukaryotic CytoskeletonTathagata ChakravarttyNo ratings yet
- MicrotubulesDocument39 pagesMicrotubulesAdnan QureshiNo ratings yet
- Cytoskeleton PRESENTATIONDocument36 pagesCytoskeleton PRESENTATIONChaudryNomiNo ratings yet
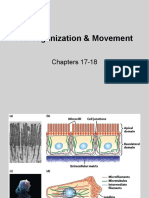
- 17&18) Cell Organization & MovementDocument28 pages17&18) Cell Organization & MovementRNo ratings yet
- Prepared By:-: Priyanka Yadav M.Sc. Life Sciences Ist SemDocument37 pagesPrepared By:-: Priyanka Yadav M.Sc. Life Sciences Ist SemleartaNo ratings yet
- Cy To SkeletonDocument21 pagesCy To SkeletonJisna JaisonNo ratings yet
- Cytoskelet ON: Dr. I Wayan Sugiritama, M.KesDocument42 pagesCytoskelet ON: Dr. I Wayan Sugiritama, M.KesPande Indra PremanaNo ratings yet
- BIOL2120 7 Cell CycleDocument44 pagesBIOL2120 7 Cell CycleHui Ka HoNo ratings yet
- Lecture 3 CytoskeletonfsDocument54 pagesLecture 3 Cytoskeletonfsaidar.seralinNo ratings yet
- The Cytoskeleton: Shareeka Joana NgilayDocument37 pagesThe Cytoskeleton: Shareeka Joana NgilayleartaNo ratings yet
- The CellDocument24 pagesThe CellTyra Joyce Revadavia100% (1)
- CytosceletonDocument41 pagesCytosceletonaidar.seralinNo ratings yet
- Cellular MovementsDocument27 pagesCellular MovementsngidizzmNo ratings yet
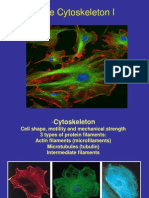
- The CytoskeletonDocument63 pagesThe CytoskeletonAmara TargaryenNo ratings yet
- Thecytoskeleton PPTDocument11 pagesThecytoskeleton PPTseenu mohapatra100% (1)
- Cytoskeleton and Cell Motility 1Document40 pagesCytoskeleton and Cell Motility 1Muhamad Farhan RLNo ratings yet
- Cell ModularmodifiedDocument292 pagesCell ModularmodifiedkofffeeeeNo ratings yet
- Lecture 15-Cell BiologyDocument36 pagesLecture 15-Cell BiologyJhon Jairo SarmientoNo ratings yet
- (B) Microfilaments: FunctionDocument2 pages(B) Microfilaments: FunctionDrbee10No ratings yet
- Mitokondria 1Document40 pagesMitokondria 1Nilam CahyaNo ratings yet
- Molecular Bioeng-2: Mrs Daramola BellstechDocument22 pagesMolecular Bioeng-2: Mrs Daramola BellstechSamuel EkpoNo ratings yet
- Notes 3.1 - CYTOPLASM PART 4Document5 pagesNotes 3.1 - CYTOPLASM PART 4Kahlen SivanNo ratings yet
- Biomolecules & Cells Biol10002 Lectures 1-9 Cell Biology Geoff Mcfadden, RM 211 Biosciences 2 (Old Botany)Document41 pagesBiomolecules & Cells Biol10002 Lectures 1-9 Cell Biology Geoff Mcfadden, RM 211 Biosciences 2 (Old Botany)waimoeNo ratings yet
- The Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellDocument9 pagesThe Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellHania GulNo ratings yet
- Human Physiology and Anatomy: LEC 01 Cells and TissuesDocument47 pagesHuman Physiology and Anatomy: LEC 01 Cells and TissuesPersonal growthNo ratings yet
- Extracellular Matrix and CytoskeletonDocument23 pagesExtracellular Matrix and CytoskeletonjoyyeesufNo ratings yet
- Platelet Production, Structure, & FunctionDocument88 pagesPlatelet Production, Structure, & FunctionphyjustineNo ratings yet
- 5 - Cytoskeleton and Cell MotilityDocument21 pages5 - Cytoskeleton and Cell Motilitywhether913No ratings yet
- CytoskeletonDocument81 pagesCytoskeletonMugaboNo ratings yet
- Cytoskeleton for Class 1Document32 pagesCytoskeleton for Class 1seerats770No ratings yet
- 2022 CytoskeletonDocument59 pages2022 CytoskeletonAwais RehmanNo ratings yet
- Cytoskeleton SNDocument3 pagesCytoskeleton SNsamy gNo ratings yet
- Cytoskeleton (1) HBHHHHGGH, HGDocument24 pagesCytoskeleton (1) HBHHHHGGH, HGrohanv4712No ratings yet
- Cell CytoskeletonDocument7 pagesCell Cytoskeletonandersonmack047No ratings yet
- 11.sitoskeleton Dan Pergerakan Sel-Dr - IsraDocument40 pages11.sitoskeleton Dan Pergerakan Sel-Dr - IsraTa RaNo ratings yet
- BY2201 EmmaC CytoskeletonIandIIDocument62 pagesBY2201 EmmaC CytoskeletonIandIIRebecca ReillyNo ratings yet
- Cytoskeletal SystemDocument28 pagesCytoskeletal Systemswarnaprabhadas07No ratings yet
- Lecture 6 - CytoskeletonDocument38 pagesLecture 6 - CytoskeletonNL DearestNo ratings yet
- From Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.From EverandFrom Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.No ratings yet
- Ch. 13-Cytoskeletal SystemsDocument18 pagesCh. 13-Cytoskeletal Systemsnkorkmaz1357No ratings yet
- Biol2120 Cell Biology: 8. Apoptosis: TheDocument28 pagesBiol2120 Cell Biology: 8. Apoptosis: TheHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationDocument49 pagesBiol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationDocument49 pagesBiol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationHui Ka HoNo ratings yet
- BIOL2120 7 Cell CycleDocument44 pagesBIOL2120 7 Cell CycleHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Signal Transduction Mechanisms: II. Messengers and ReceptorsDocument58 pagesBiol2120 Cell Biology: Signal Transduction Mechanisms: II. Messengers and ReceptorsHui Ka HoNo ratings yet
- BIOL2120 3 Transport Across MembranesDocument60 pagesBIOL2120 3 Transport Across MembranesHui Ka HoNo ratings yet
- Biol2120 Cell Biology: 9. CancerDocument28 pagesBiol2120 Cell Biology: 9. CancerHui Ka HoNo ratings yet
- Biol2120 Cell Biology: 8. Apoptosis: TheDocument28 pagesBiol2120 Cell Biology: 8. Apoptosis: TheHui Ka HoNo ratings yet
- BIOL2120 3 Transport Across MembranesDocument60 pagesBIOL2120 3 Transport Across MembranesHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Signal Transduction Mechanisms: II. Messengers and ReceptorsDocument58 pagesBiol2120 Cell Biology: Signal Transduction Mechanisms: II. Messengers and ReceptorsHui Ka HoNo ratings yet
- BIOL2120 4 Endomembrane SystemDocument47 pagesBIOL2120 4 Endomembrane SystemHui Ka HoNo ratings yet
- SQL 1: Basic Statements: Yufei TaoDocument24 pagesSQL 1: Basic Statements: Yufei TaoHui Ka HoNo ratings yet
- Yufei Tao: Department of Computer Science and Engineering Chinese University of Hong KongDocument18 pagesYufei Tao: Department of Computer Science and Engineering Chinese University of Hong KongHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Membranes: Their Structure, Function, and Chemistry 2. MembranesDocument49 pagesBiol2120 Cell Biology: Membranes: Their Structure, Function, and Chemistry 2. MembranesHui Ka HoNo ratings yet
- Rsa ProofDocument10 pagesRsa ProofHui Ka HoNo ratings yet
- Table Definition and Modification: Yufei TaoDocument13 pagesTable Definition and Modification: Yufei TaoHui Ka HoNo ratings yet
- Bosco Public School: Invitation For Virtual Fest-'Spectrum 2021'Document9 pagesBosco Public School: Invitation For Virtual Fest-'Spectrum 2021'Mehul KapoorNo ratings yet
- Data Analysis Lesson PlanDocument3 pagesData Analysis Lesson PlanBriLynn83No ratings yet
- Data Structures With C++Document169 pagesData Structures With C++Rachna BhatnagarNo ratings yet
- Deep Excavation BasementDocument8 pagesDeep Excavation BasementTan 程惠萍No ratings yet
- HTK (v.3.1) : Basic Tutorial: ContentDocument18 pagesHTK (v.3.1) : Basic Tutorial: ContentJanković MilicaNo ratings yet
- LTRT-83309 Mediant 600 and Mediant 1000 SIP User's Manual v6.4Document824 pagesLTRT-83309 Mediant 600 and Mediant 1000 SIP User's Manual v6.4kaz7878No ratings yet
- Detail Tangga Mockup PDFDocument1 pageDetail Tangga Mockup PDFanicoNo ratings yet
- Joint Angle-Based EMG Amplitude Calibration: ArticleDocument5 pagesJoint Angle-Based EMG Amplitude Calibration: ArticlekorsairNo ratings yet
- Unigraphics NX Interview Questions and AnswersDocument5 pagesUnigraphics NX Interview Questions and AnswersVivekPJNo ratings yet
- 1 BoardCompanion Physics PDFDocument61 pages1 BoardCompanion Physics PDFSrn YuvaneshNo ratings yet
- Plasma Kata LogDocument16 pagesPlasma Kata Logmurat123usNo ratings yet
- Aoc 18.5Document46 pagesAoc 18.5Sanskar PandaNo ratings yet
- Project Report On Tpms DeviceDocument12 pagesProject Report On Tpms DeviceVikas KumarNo ratings yet
- 1MA1 3F Rms 20240111Document20 pages1MA1 3F Rms 20240111jorgeivan3458No ratings yet
- Determination of Negative Permittivity and PermeabDocument6 pagesDetermination of Negative Permittivity and PermeabNewton GomesNo ratings yet
- Intel® Ethernet Converged Network Adapter XL710: Product BriefDocument4 pagesIntel® Ethernet Converged Network Adapter XL710: Product Briefmatthew_mk_au2271No ratings yet
- 73 Magazine 01 - January - 1961Document68 pages73 Magazine 01 - January - 1961viajerocanarioNo ratings yet
- Installation Manual: EnglishDocument24 pagesInstallation Manual: EnglishAntonio FavuzziNo ratings yet
- 7-7-2019 19 55 42 ErrorLogDocument4 pages7-7-2019 19 55 42 ErrorLogMarvin CustodioNo ratings yet
- QB WTDocument66 pagesQB WTSiddhi GavhaneNo ratings yet
- Cost Allocation SAP HANADocument47 pagesCost Allocation SAP HANAAmy Nguyễn100% (1)
- Sliding Mode Control Strategy For A 6 DOF Quadrotor HelicopterDocument7 pagesSliding Mode Control Strategy For A 6 DOF Quadrotor HelicopternaderjsaNo ratings yet
- Low-Power and Fast Full Adder by Exploring New and Gates: XOR XnorDocument13 pagesLow-Power and Fast Full Adder by Exploring New and Gates: XOR XnorBhargav Naidu PulugujjuNo ratings yet
- Week 1 QuizDocument20 pagesWeek 1 QuizEduardo GomezNo ratings yet
- C - Fakepathsyllabus Linear Algebra2023 2Document6 pagesC - Fakepathsyllabus Linear Algebra2023 2ywgf4q7hkkNo ratings yet
- Domain Specific Languages in Practice: International Journal of Computer Applications April 2015Document5 pagesDomain Specific Languages in Practice: International Journal of Computer Applications April 2015Radian Nata KristiantoNo ratings yet
- Ultratech Cement: Particulars Test Results Requirements ofDocument1 pageUltratech Cement: Particulars Test Results Requirements ofAshish SontakkeNo ratings yet
- HPCL SYLLABUS 2022 Chemical PediaDocument6 pagesHPCL SYLLABUS 2022 Chemical PediaRudra MishraNo ratings yet