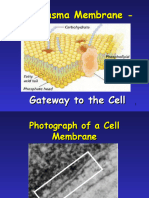
Download as pdf or txt
You might also like
- Commercial Case Study Candidate Info Pack PDFDocument29 pagesCommercial Case Study Candidate Info Pack PDFGoogxxy67% (3)
- Harley Davidson Vs Honda StrategyDocument26 pagesHarley Davidson Vs Honda StrategyKislaya Sharma33% (3)
- Cabin Crew Interview Preparation Practice Booklet Cabin Crew Excellence EbookDocument15 pagesCabin Crew Interview Preparation Practice Booklet Cabin Crew Excellence EbookLIZANNENo ratings yet
- Method Statement of Passenger Hoist Indonesia 1 Project 1-08-2018Document37 pagesMethod Statement of Passenger Hoist Indonesia 1 Project 1-08-2018Andhika Herdiawan100% (1)
- Quality Assurance For MS PipeDocument54 pagesQuality Assurance For MS PipeRam Narendra Nath100% (2)
- t5 SwapDocument19 pagest5 SwapSean Cross100% (2)
- BIOL2120 3 Transport Across MembranesDocument60 pagesBIOL2120 3 Transport Across MembranesHui Ka HoNo ratings yet
- Chapter 8 Cellular Membranes Part 2, Biols300Document46 pagesChapter 8 Cellular Membranes Part 2, Biols300mariamNo ratings yet
- Cell Membrane Transport PhysioDocument37 pagesCell Membrane Transport PhysioWandera GodfreyNo ratings yet
- Membrane TransportDocument24 pagesMembrane Transportolawandeilo123No ratings yet
- Biology NotesDocument37 pagesBiology Noteschewe cheweNo ratings yet
- Pharmacy Lecture Note Transport Across Cell Membranes Feyitimi AaDocument44 pagesPharmacy Lecture Note Transport Across Cell Membranes Feyitimi AaMarlene NiyomuhozaNo ratings yet
- BBT221 Lecture 2Document19 pagesBBT221 Lecture 2Al Sabri Bhuiyan 1812098042No ratings yet
- Cell Membranes and TransportDocument32 pagesCell Membranes and TransportTobiNo ratings yet
- 2014 Molecular Kinetic of DrugsDocument49 pages2014 Molecular Kinetic of DrugsDarry AprilioNo ratings yet
- Botafun Biological MembranesDocument9 pagesBotafun Biological Membraneslily remNo ratings yet
- Slides: Slideshow View. Use The Buttons On TheDocument46 pagesSlides: Slideshow View. Use The Buttons On TheAna Luísa FerreiraNo ratings yet
- Transport Across MembranesDocument61 pagesTransport Across MembranesAmara TargaryenNo ratings yet
- Life Science Notes Q3 Week 3Document34 pagesLife Science Notes Q3 Week 3Juan Miguel Dela CruzNo ratings yet
- L3The Cell Structure and Functions PDFDocument45 pagesL3The Cell Structure and Functions PDFVince Martin ManaigNo ratings yet
- Lecture 2 Transport of Substances Across The Cell MembraneDocument24 pagesLecture 2 Transport of Substances Across The Cell MembraneEzeudu SomtoNo ratings yet
- Membranes and Transport: SlidesDocument31 pagesMembranes and Transport: SlidesZohal HayeriNo ratings yet
- CH 3Document37 pagesCH 3Viraat SinghNo ratings yet
- Chapter 7Document21 pagesChapter 7EhazNo ratings yet
- Physiology U-2 Transport Across The Plasma MembraneDocument48 pagesPhysiology U-2 Transport Across The Plasma Membranesinte beyuNo ratings yet
- WEEK 7 TransportDocument49 pagesWEEK 7 Transportorioluna00No ratings yet
- The Cellular Level of Organization: - Basic, Living, Structural and Functional Unit of The BodyDocument76 pagesThe Cellular Level of Organization: - Basic, Living, Structural and Functional Unit of The BodyAyush KcNo ratings yet
- FHCG 2 - Cell Organisation CommunicationDocument68 pagesFHCG 2 - Cell Organisation CommunicationLeena AkhtarNo ratings yet
- Cell Lecture 5-1Document45 pagesCell Lecture 5-1gracechirangoNo ratings yet
- Membrane Transport System - BiochemDocument25 pagesMembrane Transport System - BiochemShreepriya KhuranaNo ratings yet
- Membrane - FunctionDocument4 pagesMembrane - Functionphuong anhNo ratings yet
- Membrane - FunctionDocument4 pagesMembrane - Functionphuong anhNo ratings yet
- Chapter 5 BioDocument12 pagesChapter 5 BioGeorges SarkisNo ratings yet
- Lesson 5. Cell Transport NOTESDocument76 pagesLesson 5. Cell Transport NOTESPaolo RapanizaNo ratings yet
- Transport Across The Cell Membrane - OdpDocument67 pagesTransport Across The Cell Membrane - OdpSilver CypressNo ratings yet
- Topic 1.4Document17 pagesTopic 1.4Fatima ZaidiNo ratings yet
- Plasma Membrane Structure and FunctionDocument50 pagesPlasma Membrane Structure and Functiondhea wirantiNo ratings yet
- Topic 4Document8 pagesTopic 4angelinatianranNo ratings yet
- Cell Structure and FunctionsDocument10 pagesCell Structure and FunctionsQsh SinoroNo ratings yet
- General Physiology Transport Across The Cell Membrane: By: Dr. Hamza Al-TradDocument19 pagesGeneral Physiology Transport Across The Cell Membrane: By: Dr. Hamza Al-TradibrahimNo ratings yet
- 10 - Transportation and Lipid Bilayer - 2020Document29 pages10 - Transportation and Lipid Bilayer - 2020tireddoodleNo ratings yet
- Transpor Trans MembranDocument48 pagesTranspor Trans Membrananandaputriaisyah19No ratings yet
- Plasma MembraneDocument66 pagesPlasma Membranenurul atika100% (2)
- Cell Membrane - Corrected by SheenaDocument46 pagesCell Membrane - Corrected by SheenaJasmine YiNo ratings yet
- CH5 Membrane TraffickingDocument48 pagesCH5 Membrane TraffickingAhmed MohamadNo ratings yet
- 1.4 Membrane TransportDocument41 pages1.4 Membrane TransportZeineb AbbasNo ratings yet
- Transport Across Cell MembraneDocument40 pagesTransport Across Cell MembraneMohamed NourNo ratings yet
- CytologyDocument126 pagesCytologyKamal AhmedNo ratings yet
- Bio Chapter 3Document33 pagesBio Chapter 3Athirah BadrulshamNo ratings yet
- L3 Cell Membrane and Transport 2Document23 pagesL3 Cell Membrane and Transport 2sampsonsoo17No ratings yet
- Membrane Transport ABDocument38 pagesMembrane Transport ABdj77djcqy7No ratings yet
- Plasma MembraneDocument60 pagesPlasma MembraneJinky PeraltaNo ratings yet
- Lec 3 Transport Across Cell MembraneDocument22 pagesLec 3 Transport Across Cell MembraneHafiz NaveedNo ratings yet
- Transport MechanismDocument105 pagesTransport MechanismMAY FULGENCIONo ratings yet
- Kognity Notes Bio B2Document20 pagesKognity Notes Bio B2eybasco.2001No ratings yet
- San Pedro College: Senior High School DepartmentDocument54 pagesSan Pedro College: Senior High School DepartmentKrisha Diane FunaNo ratings yet
- 1 Cellular Transport VidDocument57 pages1 Cellular Transport Vidchizy bananaNo ratings yet
- Cell MembraneDocument60 pagesCell Membranejjcy020529No ratings yet
- Ch11 Trans Membrane Transport of Ions and Small MoleculeDocument113 pagesCh11 Trans Membrane Transport of Ions and Small MoleculeamuthukumaranNo ratings yet
- CH - 6, Passive TransportDocument43 pagesCH - 6, Passive TransportTony StarkNo ratings yet
- Structure of Cell Membrane Cell TransportDocument4 pagesStructure of Cell Membrane Cell TransportMarjhon playsmcpe787No ratings yet
- Transport Function: of CellDocument41 pagesTransport Function: of CellDr. R. PeriasamyNo ratings yet
- (W3) Cellular Level of OrganizationDocument4 pages(W3) Cellular Level of OrganizationFRANCES DEANNE ALABADONo ratings yet
- Lecture On Membrane Transport by Dr. Mudassar Ali Roomi.Document37 pagesLecture On Membrane Transport by Dr. Mudassar Ali Roomi.Mudassar Roomi100% (1)
- Biol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationDocument49 pagesBiol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationDocument49 pagesBiol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationHui Ka HoNo ratings yet
- Biol2120 Cell Biology: 8. Apoptosis: TheDocument28 pagesBiol2120 Cell Biology: 8. Apoptosis: TheHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Signal Transduction Mechanisms: II. Messengers and ReceptorsDocument58 pagesBiol2120 Cell Biology: Signal Transduction Mechanisms: II. Messengers and ReceptorsHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationDocument49 pagesBiol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationHui Ka HoNo ratings yet
- BIOL2120 7 Cell CycleDocument44 pagesBIOL2120 7 Cell CycleHui Ka HoNo ratings yet
- Biol2120 Cell Biology: 9. CancerDocument28 pagesBiol2120 Cell Biology: 9. CancerHui Ka HoNo ratings yet
- Biol2120 Cell Biology: 8. Apoptosis: TheDocument28 pagesBiol2120 Cell Biology: 8. Apoptosis: TheHui Ka HoNo ratings yet
- BIOL2120 3 Transport Across MembranesDocument60 pagesBIOL2120 3 Transport Across MembranesHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Signal Transduction Mechanisms: II. Messengers and ReceptorsDocument58 pagesBiol2120 Cell Biology: Signal Transduction Mechanisms: II. Messengers and ReceptorsHui Ka HoNo ratings yet
- BIOL2120 4 Endomembrane SystemDocument47 pagesBIOL2120 4 Endomembrane SystemHui Ka HoNo ratings yet
- SQL 1: Basic Statements: Yufei TaoDocument24 pagesSQL 1: Basic Statements: Yufei TaoHui Ka HoNo ratings yet
- Yufei Tao: Department of Computer Science and Engineering Chinese University of Hong KongDocument18 pagesYufei Tao: Department of Computer Science and Engineering Chinese University of Hong KongHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Membranes: Their Structure, Function, and Chemistry 2. MembranesDocument49 pagesBiol2120 Cell Biology: Membranes: Their Structure, Function, and Chemistry 2. MembranesHui Ka HoNo ratings yet
- Rsa ProofDocument10 pagesRsa ProofHui Ka HoNo ratings yet
- Table Definition and Modification: Yufei TaoDocument13 pagesTable Definition and Modification: Yufei TaoHui Ka HoNo ratings yet
- The Boeing 737 Max Saga Automating FailureDocument2 pagesThe Boeing 737 Max Saga Automating FailureGökçen AkmaNo ratings yet
- BarillaDocument24 pagesBarilladkotulski12No ratings yet
- Referensi Unaccompanied 1Document38 pagesReferensi Unaccompanied 1Tegar Dwi PriambodoNo ratings yet
- Civacon Manifold Intallation and Parts Breakdowns 27 PagesDocument2 pagesCivacon Manifold Intallation and Parts Breakdowns 27 PagesMohd FairusNo ratings yet
- OHE Masts PDFDocument39 pagesOHE Masts PDFManvendra SinghNo ratings yet
- Lec 1 - SuperelevationDocument24 pagesLec 1 - SuperelevationMohamedNo ratings yet
- Expansion Bom 10-2022 Option 7 ApprovedDocument11 pagesExpansion Bom 10-2022 Option 7 ApprovedSwazelle DianeNo ratings yet
- ITA WG6 - 2017 FINAL EditionDocument66 pagesITA WG6 - 2017 FINAL EditionNicolas LozanoNo ratings yet
- Caf News 80Document56 pagesCaf News 80Blue Lou100% (1)
- Signal Timing and DesignDocument12 pagesSignal Timing and DesignVirtual RealityNo ratings yet
- Transport Department. Govt of NCT of Delhi: Form 1-A (See Rules 5 (1), (3), 7,10 (A), 14 (D), and 18 (D) )Document1 pageTransport Department. Govt of NCT of Delhi: Form 1-A (See Rules 5 (1), (3), 7,10 (A), 14 (D), and 18 (D) )Rãhûl SâïñíNo ratings yet
- Gondola Inspection ChecklistDocument1 pageGondola Inspection ChecklistPaul Adrian CasNo ratings yet
- B2B Collaboration Flowchart: Electronic MallDocument1 pageB2B Collaboration Flowchart: Electronic MallanggonoutomoNo ratings yet
- GP Rating English LeafletDocument2 pagesGP Rating English LeafletSunik KatochNo ratings yet
- General Motors CompanyDocument8 pagesGeneral Motors CompanyNeha Soningra100% (1)
- OFPER 1 Application For Seagoing AppointmentDocument6 pagesOFPER 1 Application For Seagoing AppointmentMelden Roy VisandeNo ratings yet
- 05-13-14 PC FINAL Draft PacketDocument926 pages05-13-14 PC FINAL Draft PacketPlanningCommissionNo ratings yet
- Sprinter P-XP en 0314Document12 pagesSprinter P-XP en 0314Mohamed Ali Mahmoud SalemNo ratings yet
- English HAZET 3488 - 8 FordDocument13 pagesEnglish HAZET 3488 - 8 FordkallatisNo ratings yet
- VIN - How To Read A Subar..Document18 pagesVIN - How To Read A Subar..darfri100% (1)
- New 001 2014 HYDocument4 pagesNew 001 2014 HYРома БлошенкоNo ratings yet
- Revista de TransmisionesDocument68 pagesRevista de TransmisionesTransmisiones Automáticas Chepe100% (4)
- A Transportation Ontology For Global City Indicators (ISO 37120) 1.0Document57 pagesA Transportation Ontology For Global City Indicators (ISO 37120) 1.0Csaba HitterNo ratings yet
- Progress Tracker-HVAC & BVDocument11 pagesProgress Tracker-HVAC & BVInfant VelladuraiNo ratings yet