Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5823)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Homeostasis Lab ActivitiesDocument6 pagesHomeostasis Lab Activitiesapi-34456850050% (2)
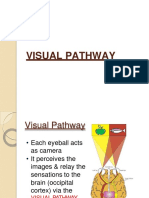
- Visual PathwayDocument24 pagesVisual PathwayAkshara Eye FoundationNo ratings yet
- 1 PBDocument9 pages1 PBtruckerpunkNo ratings yet
- 96 - 2013 XiuDocument6 pages96 - 2013 XiutruckerpunkNo ratings yet
- Johnson 1954Document3 pagesJohnson 1954truckerpunkNo ratings yet
- Miranda 2016Document6 pagesMiranda 2016truckerpunkNo ratings yet
- ExperimentDocument5 pagesExperimenttruckerpunkNo ratings yet
- Wagh 832015 Arr B 19936Document15 pagesWagh 832015 Arr B 19936truckerpunkNo ratings yet
- Ohaus MB200 ManualDocument37 pagesOhaus MB200 ManualtruckerpunkNo ratings yet
- Repurposing Tempeh Fermentation: A Promising Protein Source Using Food Residues and Edible Filamentous FungiDocument6 pagesRepurposing Tempeh Fermentation: A Promising Protein Source Using Food Residues and Edible Filamentous FungitruckerpunkNo ratings yet
- Fermentation 09 00291 v2Document15 pagesFermentation 09 00291 v2truckerpunkNo ratings yet
- Chemical Actinometry: Using O-Nitrobenzaldehyde To Measure Light Intensity in Photochemical ExperimentsDocument3 pagesChemical Actinometry: Using O-Nitrobenzaldehyde To Measure Light Intensity in Photochemical ExperimentstruckerpunkNo ratings yet
- Feng 2005 GrowthoflacticaciDocument9 pagesFeng 2005 GrowthoflacticacitruckerpunkNo ratings yet
- Pain HistoricaloverviewDocument17 pagesPain HistoricaloverviewtruckerpunkNo ratings yet
- A Collection of Experiments For Teaching PhotochemistryDocument40 pagesA Collection of Experiments For Teaching PhotochemistrytruckerpunkNo ratings yet
- Energetic PolymersDocument13 pagesEnergetic PolymerstruckerpunkNo ratings yet
- A Brief History of Opiates, Opioid Peptides, and Opioid ReceptorsDocument3 pagesA Brief History of Opiates, Opioid Peptides, and Opioid ReceptorstruckerpunkNo ratings yet
- Development of Supercapacitors Based On Conducting Polymers and Its Composites With GrapheneDocument36 pagesDevelopment of Supercapacitors Based On Conducting Polymers and Its Composites With GraphenetruckerpunkNo ratings yet
- 15-Organic Synthesis PDFDocument30 pages15-Organic Synthesis PDFtruckerpunkNo ratings yet
- Azatrioxa (8) CirculeneDocument79 pagesAzatrioxa (8) CirculenetruckerpunkNo ratings yet
- General Biology q1 w2 Mod2Document40 pagesGeneral Biology q1 w2 Mod2Karen Mae CastilloNo ratings yet
- Physiology of Male ReproductionDocument19 pagesPhysiology of Male Reproductionalok nayakNo ratings yet
- Anatomy Question BankDocument13 pagesAnatomy Question BankMahnaz MushtaqNo ratings yet
- EMT Static and Dynamic Posture LectureDocument9 pagesEMT Static and Dynamic Posture LectureBennat RajNo ratings yet
- Outline: Outline:: BI/CH 422/622Document9 pagesOutline: Outline:: BI/CH 422/622Khadija JaraNo ratings yet
- Bio IA Working DocumentDocument11 pagesBio IA Working DocumentflorryNo ratings yet
- Heart AttackDocument4 pagesHeart Attackards mcaffeeNo ratings yet
- Gall Bladder & Pancreas HistologyDocument48 pagesGall Bladder & Pancreas HistologyGood BoyNo ratings yet
- Kent Marianito-WPS OfficeDocument12 pagesKent Marianito-WPS OfficeKent Andojar MarianitoNo ratings yet
- Anatomy of PharynxDocument34 pagesAnatomy of PharynxNasr HudaNo ratings yet
- Strength For WarfightersDocument12 pagesStrength For Warfightersbeltran230111No ratings yet
- Biochem LEC-Activity 1Document2 pagesBiochem LEC-Activity 1PATRICIA DIANNE EHONGNo ratings yet
- Review - Animal Motor SystemsDocument4 pagesReview - Animal Motor SystemskjhNo ratings yet
- SCIENCE-9-Q1-Week1-MELC05-ASP-Calabucal, Ma. Belinda - Docx (2) - Ma. BELINDA CALABUCALDocument9 pagesSCIENCE-9-Q1-Week1-MELC05-ASP-Calabucal, Ma. Belinda - Docx (2) - Ma. BELINDA CALABUCALNOVA LESLIE AGAPAYNo ratings yet
- Comprehensive P. Health AssessmentDocument3 pagesComprehensive P. Health AssessmentCristoper BodionganNo ratings yet
- Contents of All Volumes: Natural Products Structural Diversity-I Secondary Metabolites: Organization and BiosynthesisDocument1,327 pagesContents of All Volumes: Natural Products Structural Diversity-I Secondary Metabolites: Organization and BiosynthesisMarcio BragaNo ratings yet
- Medical CertificateDocument5 pagesMedical CertificatePrasanna VenkatNo ratings yet
- Lecture Bee Hives and Their Description, Bee Pasturage, Bee Foraging, Behaviour and CommunicationDocument19 pagesLecture Bee Hives and Their Description, Bee Pasturage, Bee Foraging, Behaviour and CommunicationVIVEK SUTARNo ratings yet
- PhotosynthesisDocument17 pagesPhotosynthesisAnirban DeNo ratings yet
- PDF Stress Physiology of Woody Plants 1St Edition Wenhao Dai Ebook Full ChapterDocument53 pagesPDF Stress Physiology of Woody Plants 1St Edition Wenhao Dai Ebook Full Chapterjennifer.nye440100% (1)
- Semi-Detailed Lesson Plan in Science I.Objective: V Fomuh8D3N7WDocument4 pagesSemi-Detailed Lesson Plan in Science I.Objective: V Fomuh8D3N7WTintin Dimalanta LacanlaleNo ratings yet
- Park Ridge School of Montessori IncorporatedDocument53 pagesPark Ridge School of Montessori IncorporatedJustine DakissNo ratings yet
- BiosensorsDocument28 pagesBiosensorsTanvi SharmaNo ratings yet
- Human Embryonic Development - WikipediaDocument13 pagesHuman Embryonic Development - WikipediaHassan UsmaniNo ratings yet
- Heredity and Evolution Notes. SSC Class 10Document24 pagesHeredity and Evolution Notes. SSC Class 10ONI SAITAMA100% (1)
- Lecture 13 - Conditioned & Unconditioned ReflexesDocument40 pagesLecture 13 - Conditioned & Unconditioned Reflexesסנאבל מוסאNo ratings yet
- Physiological Psychology - FinalsDocument40 pagesPhysiological Psychology - Finalsmx2r6pv778No ratings yet
- Session #43 SAS - AnaPhy (Lab) (Agdana, Nicole Ken)Document4 pagesSession #43 SAS - AnaPhy (Lab) (Agdana, Nicole Ken)Nicole Ken AgdanaNo ratings yet