Download as pdf or txt
You might also like
- Freud Bose CorrespondenceDocument17 pagesFreud Bose CorrespondenceSiddhi JangidNo ratings yet
- Thiabendazole (WHO Food Additives Series 39)Document7 pagesThiabendazole (WHO Food Additives Series 39)hellen86150No ratings yet
- The Effects of Nutrition On Unconjugated Plasma Bilirubin Concentrations in Squirrel MonkeysDocument7 pagesThe Effects of Nutrition On Unconjugated Plasma Bilirubin Concentrations in Squirrel MonkeysNadzhin15No ratings yet
- Norgate 2021Document8 pagesNorgate 2021Widya WibelNo ratings yet
- Reduced Diabetogenic Effect of Streptozotocin in Rats Previously Adapted To A High-Protein, Carbohydrate-Free DietDocument6 pagesReduced Diabetogenic Effect of Streptozotocin in Rats Previously Adapted To A High-Protein, Carbohydrate-Free DietPaul SimononNo ratings yet
- Effects of Pair-Feeding and Growth Hormone Treatment On Obese Transgenic RatsDocument5 pagesEffects of Pair-Feeding and Growth Hormone Treatment On Obese Transgenic RatsThanh LeNo ratings yet
- Induction of Diabetes by Streptozotocin in RatsDocument5 pagesInduction of Diabetes by Streptozotocin in RatsJusty GuavaNo ratings yet
- Akbarzadeh 2007 Induksi STZ Diabetic PDFDocument5 pagesAkbarzadeh 2007 Induksi STZ Diabetic PDFPutri MauraNo ratings yet
- Chron y Glutamina Denhond1999Document5 pagesChron y Glutamina Denhond1999paulabagnaroliNo ratings yet
- InsulinDocument6 pagesInsulinAuthor Nauman ShadNo ratings yet
- Original Article Evidence of Anti-Obesity Effects of The Pomegranate Leaf Extract in High-Fat Diet Induced Obese MiceDocument7 pagesOriginal Article Evidence of Anti-Obesity Effects of The Pomegranate Leaf Extract in High-Fat Diet Induced Obese MiceAisy Savira anizarNo ratings yet
- Urine Dipsticks Are Not Accurate For Detecting Mild KetosisDocument8 pagesUrine Dipsticks Are Not Accurate For Detecting Mild KetosisAmbar Sofía MieresNo ratings yet
- Effects of Diet Restriction On Life Span and Age-Related Changes in DogsDocument6 pagesEffects of Diet Restriction On Life Span and Age-Related Changes in DogsLiana OpriţăNo ratings yet
- Xu 2010 Carbohydrate-To-Fat Ratio Affects Food Intake and Body Weight in Wistar RatsDocument6 pagesXu 2010 Carbohydrate-To-Fat Ratio Affects Food Intake and Body Weight in Wistar RatsPaul SimononNo ratings yet
- Assessment of Four Experimental Models of Hyperlipidemia: Research NoteDocument6 pagesAssessment of Four Experimental Models of Hyperlipidemia: Research NoteMuhammad AwaisNo ratings yet
- Journal of Nutrition College, Volume 3, Nomor 4, Tahun 2014, Halaman 506-516Document8 pagesJournal of Nutrition College, Volume 3, Nomor 4, Tahun 2014, Halaman 506-516selvi aklailia rosaNo ratings yet
- Antihypertensive and Metabolic Effects of Whole Maitake Mushroom Powder and Its Fractions in Two Rat StrainsDocument8 pagesAntihypertensive and Metabolic Effects of Whole Maitake Mushroom Powder and Its Fractions in Two Rat StrainsAryo WirantoNo ratings yet
- Hypergammaglobulinemia and Albumin Synthesis in The RabbitDocument2 pagesHypergammaglobulinemia and Albumin Synthesis in The RabbityanuararipratamaNo ratings yet
- A Case of A Colocolic Intussusception in A HorseDocument5 pagesA Case of A Colocolic Intussusception in A HorseTiago PaixãoNo ratings yet
- Brown 1998Document9 pagesBrown 1998christian.sincheNo ratings yet
- Anthropometrical Parameters and Markers of Obesity in RatsDocument9 pagesAnthropometrical Parameters and Markers of Obesity in RatsSammer BurgosNo ratings yet
- Acido FlavaspidicoDocument7 pagesAcido FlavaspidicoKarlo Josué Sanchez RiveraNo ratings yet
- Art. DanazolDocument9 pagesArt. DanazolmrintraNo ratings yet
- Graham 1994Document4 pagesGraham 1994sergio.lopez07No ratings yet
- Screenshot 2021-10-27 at 18.10.44Document9 pagesScreenshot 2021-10-27 at 18.10.44elok_88No ratings yet
- Focal Single Intravenous Aminonucleoside: Following Dose PuromycinDocument7 pagesFocal Single Intravenous Aminonucleoside: Following Dose PuromycinFauziana NurhanisahNo ratings yet
- 1472 6882 2 1 PDFDocument7 pages1472 6882 2 1 PDFThiago NunesNo ratings yet
- Of in Vitro.: Effects Hormones On Post-Implantation Mouse EmbryosDocument7 pagesOf in Vitro.: Effects Hormones On Post-Implantation Mouse EmbryossunNo ratings yet
- Research Article: A High Protein Diet Has No Harmful Effects: A One-Year Crossover Study in Resistance-Trained MalesDocument6 pagesResearch Article: A High Protein Diet Has No Harmful Effects: A One-Year Crossover Study in Resistance-Trained Malessiti afifahNo ratings yet
- Edr2010 289614Document10 pagesEdr2010 289614FawwaziNo ratings yet
- Ginger PublishedDocument8 pagesGinger PublishedFatimoh AbdulsalamNo ratings yet
- Jcinvest00248 0177 PDFDocument11 pagesJcinvest00248 0177 PDFluis morales garzaNo ratings yet
- Grauer, GF, Et Al., J Vet Intern Med, 2002.Document1 pageGrauer, GF, Et Al., J Vet Intern Med, 2002.Yanie Ummu ArsylaNo ratings yet
- Sciencedirect: Caloric Dose-Responsive Genes in Blood Cells Differentiate The Metabolic Status of Obese MenDocument10 pagesSciencedirect: Caloric Dose-Responsive Genes in Blood Cells Differentiate The Metabolic Status of Obese MenWa Ode Umrawati LatifNo ratings yet
- Comparison Congenital Hyperbilirubinaemia: of For Nonobstructive NonhaemolyticDocument5 pagesComparison Congenital Hyperbilirubinaemia: of For Nonobstructive NonhaemolyticTulus JavaNo ratings yet
- Lead Intoxication in A Puppy - A Case Report: E. S, J. P - G, I. K, B. P, B. A, Z. Š - PDocument2 pagesLead Intoxication in A Puppy - A Case Report: E. S, J. P - G, I. K, B. P, B. A, Z. Š - PMauricio RíosNo ratings yet
- Shin - Hypocholesterolemic Effect of Daily Fisetin Supplementation in High Fat Fed Sprague-Dawley Rats - 2013Document7 pagesShin - Hypocholesterolemic Effect of Daily Fisetin Supplementation in High Fat Fed Sprague-Dawley Rats - 2013thrNo ratings yet
- Metabolism: Clinical and ExperimentalDocument3 pagesMetabolism: Clinical and ExperimentalDawn KentNo ratings yet
- 1 s2.0 S0022030214005190 MainDocument11 pages1 s2.0 S0022030214005190 MainsiskaNo ratings yet
- Toxicological, Pathological, Teratological Studies in Animals With CephradineDocument4 pagesToxicological, Pathological, Teratological Studies in Animals With CephradinePharmacistNo ratings yet
- Aumento IGF-1 Gatos Não Diabéticos 2024Document5 pagesAumento IGF-1 Gatos Não Diabéticos 2024Lucas XavierNo ratings yet
- Effects of Aged Black Garlic Extract On Lipid Improvement in Rats Fed With High Fat-Cholesterol DietDocument9 pagesEffects of Aged Black Garlic Extract On Lipid Improvement in Rats Fed With High Fat-Cholesterol DietAnisa LestariNo ratings yet
- Estudio CerdosDocument9 pagesEstudio CerdosAniuski92No ratings yet
- Meta-Analysis of The Effects of Soy Protein Intake On Serum LipidsDocument7 pagesMeta-Analysis of The Effects of Soy Protein Intake On Serum Lipidssri novalinaNo ratings yet
- Perez Gutierrez2010Document8 pagesPerez Gutierrez2010Guilherme VarellaNo ratings yet
- In Vivo Sorbus Decora: Original ArticleDocument8 pagesIn Vivo Sorbus Decora: Original ArticleSophea Ananoria MissionaNo ratings yet
- Effect of Concentrate Supplementation On PDFDocument6 pagesEffect of Concentrate Supplementation On PDFJaks RipperNo ratings yet
- Response of C57Bl/6 Mice To A Carbohydrate-Free Diet: Research Open AccessDocument6 pagesResponse of C57Bl/6 Mice To A Carbohydrate-Free Diet: Research Open AccesspopopioNo ratings yet
- Admin,+2 +2572-Fix+feb2020Document6 pagesAdmin,+2 +2572-Fix+feb2020Rizka Uda'aNo ratings yet
- A C A D e M I C S C I e N C e SDocument3 pagesA C A D e M I C S C I e N C e SM Rasya SanimanNo ratings yet
- Long-Term Feed Intake Regulation in Sheep Is Mediated by Opioid ReceptorsDocument9 pagesLong-Term Feed Intake Regulation in Sheep Is Mediated by Opioid Receptorsjee_37No ratings yet
- Track 3 Eating Patterns and Behaviour P63Document15 pagesTrack 3 Eating Patterns and Behaviour P63Yuriko AndreNo ratings yet
- Toward Improved Management of NIDDM: A Randomized, Controlled, Pilot Intervention Using A Lowfat, Vegetarian DietDocument5 pagesToward Improved Management of NIDDM: A Randomized, Controlled, Pilot Intervention Using A Lowfat, Vegetarian DietChen Inn TanNo ratings yet
- Nutrition Therapy in Stroke - Prof. HertantoDocument32 pagesNutrition Therapy in Stroke - Prof. HertantoMuhammad FirdaudNo ratings yet
- Referensi PentingDocument9 pagesReferensi PentingAttaufiq IrawanNo ratings yet
- Islet Cell Aislamiento en Experimental D, L-Etionina Pancreatitis en Perro1-S2.0-0022480485900381-MainDocument9 pagesIslet Cell Aislamiento en Experimental D, L-Etionina Pancreatitis en Perro1-S2.0-0022480485900381-MainCarlos VargasNo ratings yet
- Alternate-Day Fasting in Nonobese Subjects: Effects On Body Weight, Body Composition, and Energy MetabolismDocument5 pagesAlternate-Day Fasting in Nonobese Subjects: Effects On Body Weight, Body Composition, and Energy MetabolismCem Tunaboylu (Student)No ratings yet
- Research Article: Improving The Reliability and Utility of Streptozotocin-Induced Rat Diabetic ModelDocument15 pagesResearch Article: Improving The Reliability and Utility of Streptozotocin-Induced Rat Diabetic ModelSipend AnatomiNo ratings yet
- The Effect of The Sodium-Glucose Cotransporter Type-2 Inhibitor Dapagliflozin On Glomerular Filtration Rate in Healthy CatsDocument6 pagesThe Effect of The Sodium-Glucose Cotransporter Type-2 Inhibitor Dapagliflozin On Glomerular Filtration Rate in Healthy CatsJohn QuickNo ratings yet
- Complementary and Alternative Medical Lab Testing Part 18: PsychiatryFrom EverandComplementary and Alternative Medical Lab Testing Part 18: PsychiatryRating: 5 out of 5 stars5/5 (1)
- Complementary and Alternative Medical Lab Testing Part 6: Liver and GallbladderFrom EverandComplementary and Alternative Medical Lab Testing Part 6: Liver and GallbladderNo ratings yet
- Chapter 7 - Morphing Skin Foams - 2018 - Morphing Wing TechnologiesDocument24 pagesChapter 7 - Morphing Skin Foams - 2018 - Morphing Wing TechnologiesRichardNo ratings yet
- Exercise 4:: Organic Components: CarbohydratesDocument20 pagesExercise 4:: Organic Components: CarbohydratespikachuzingungaNo ratings yet
- International StandardDocument8 pagesInternational Standardnazrul islamNo ratings yet
- Genetics and EugenicsDocument518 pagesGenetics and EugenicsTapuwa ChizunzaNo ratings yet
- Major Trauma Fellowship Senior Fellow Job Description: VisionDocument10 pagesMajor Trauma Fellowship Senior Fellow Job Description: VisionM LubisNo ratings yet
- Pentavitin - DSMDocument3 pagesPentavitin - DSMRnD Roi SuryaNo ratings yet
- Final DraftDocument37 pagesFinal DraftMorareng MariettaNo ratings yet
- B 17Document15 pagesB 17Rochdi SahliNo ratings yet
- Module 2 Project Gap Analysis Strategy For Team #3Document20 pagesModule 2 Project Gap Analysis Strategy For Team #3Mary A. Dillard100% (2)
- Master Circular 16 Compassionate GroundDocument13 pagesMaster Circular 16 Compassionate GroundSHANMUGA VADIVEL SUNDARARAJANNo ratings yet
- PC3SH21YFZBFDocument15 pagesPC3SH21YFZBFkrimoNo ratings yet
- Fresh and Simple PDFDocument3 pagesFresh and Simple PDFthivashnee padayacheeNo ratings yet
- Exercise 1. You Have Been Given A Form Which Shows The Details of Technical Jobs Carried OutDocument2 pagesExercise 1. You Have Been Given A Form Which Shows The Details of Technical Jobs Carried OutNgo Gia Huy (K15 HCM)No ratings yet
- Printable Cosmetology LawbookDocument42 pagesPrintable Cosmetology LawbookCaro TNo ratings yet
- SDM-v1 0Document32 pagesSDM-v1 0Franz NussmannNo ratings yet
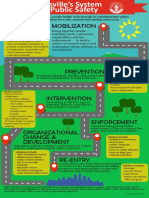
- Reimagining Public SafetyDocument1 pageReimagining Public SafetyWAVE 3 NewsNo ratings yet
- TLE Cookery10 Q4 W3 V4Document7 pagesTLE Cookery10 Q4 W3 V4Maria Rose Tariga Aquino100% (2)
- 2007-Current MaxxForce Engine Breakout Harness Reference (EGES545)Document14 pages2007-Current MaxxForce Engine Breakout Harness Reference (EGES545)Enrrique Lara100% (1)
- CreamsDocument17 pagesCreamsSolomonNo ratings yet
- Defence STD 05-50 Part 29Document14 pagesDefence STD 05-50 Part 29goodguymumbaiNo ratings yet
- DRRR2324 Q4 DRRDocument7 pagesDRRR2324 Q4 DRRh4gtg7n55gNo ratings yet
- 1A Innledning - EPS - Flyer - 2 - Jan - VaslestadDocument4 pages1A Innledning - EPS - Flyer - 2 - Jan - VaslestadNCS40 Trương Quốc BảoNo ratings yet
- Consol P 200heDocument2 pagesConsol P 200heimmanuel lumbantobingNo ratings yet
- ISC BiotechnologyDocument6 pagesISC BiotechnologySamikshaNo ratings yet
- Calibration of Dosimeters Used in Radiation TherapyDocument122 pagesCalibration of Dosimeters Used in Radiation TherapySOHON SINHA MAHAPATRANo ratings yet
- Activity 1: Measuring The Volume of Irregularly Shaped ObjectsDocument4 pagesActivity 1: Measuring The Volume of Irregularly Shaped ObjectsBamPanggatNo ratings yet
- Literature of Exile in Goli Taraqqi's Short StoriesDocument23 pagesLiterature of Exile in Goli Taraqqi's Short StoriesmarvehNo ratings yet
- Olea Europaea A Phyto-Pharmacological ReviewDocument5 pagesOlea Europaea A Phyto-Pharmacological ReviewyigalbyNo ratings yet
- Makro Chef Ingredients For Your PassionDocument62 pagesMakro Chef Ingredients For Your PassionCarina LopesNo ratings yet