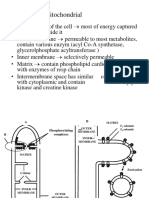
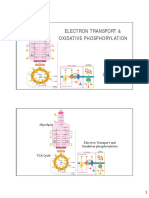
Download as pdf or txt
You might also like
- This Study Resource Was: Worksheet DDocument8 pagesThis Study Resource Was: Worksheet DBetty CooperNo ratings yet
- Electron Transport ChainDocument37 pagesElectron Transport ChainFan SmithNo ratings yet
- Gayatree PaniDocument23 pagesGayatree PaniGobindaSahuNo ratings yet
- Lehninger Principles of Biochemistry PDFDocument37 pagesLehninger Principles of Biochemistry PDFmonitamiftah100% (1)
- Electrontransportchain 151027073626 Lva1 App6892Document29 pagesElectrontransportchain 151027073626 Lva1 App6892GobindaSahuNo ratings yet
- Electron Transport ChainDocument15 pagesElectron Transport Chainvanshdeep SinghNo ratings yet
- Electron Transport ChainDocument22 pagesElectron Transport Chainbluegreenalga100% (2)
- Respiratory Chain & Oxidative PhosphorylationDocument57 pagesRespiratory Chain & Oxidative PhosphorylationHanifa AffianiNo ratings yet
- Lecture 6: Part 1 Oxidative Phosphorylation and ETC: Produced by Samira Aili Presented by Alvaro GarciaDocument23 pagesLecture 6: Part 1 Oxidative Phosphorylation and ETC: Produced by Samira Aili Presented by Alvaro Garciakristal elias100% (1)
- BBC2 K27,28RCDocument42 pagesBBC2 K27,28RCMarieta RitongaNo ratings yet
- Blue and Yellow Neon Retro Pixel Art Game Night Game Presentation - 20240124 - 211001 - 0000Document56 pagesBlue and Yellow Neon Retro Pixel Art Game Night Game Presentation - 20240124 - 211001 - 00007 Jazmin Jhem T. RullanNo ratings yet
- Electron Transport and Oxidative PhosphorylationDocument34 pagesElectron Transport and Oxidative PhosphorylationAbeWatanabeNo ratings yet
- Lecture 03 PDFDocument4 pagesLecture 03 PDFRana AmjadNo ratings yet
- Lecture 03 PDFDocument4 pagesLecture 03 PDFMuhammad TariqNo ratings yet
- Oxidative Phosphorylation 2022 PDFDocument34 pagesOxidative Phosphorylation 2022 PDFSandy IkbarNo ratings yet
- Electron Transport ch14Document21 pagesElectron Transport ch14tania.delafuenteNo ratings yet
- Cellular Respiration StationsDocument33 pagesCellular Respiration Stationsakshyta gantanNo ratings yet
- ELECTRON-TRANSPORT-CHAIN-STRYERDocument47 pagesELECTRON-TRANSPORT-CHAIN-STRYERAngelikaOdimerNo ratings yet
- Electron Transport ChainDocument4 pagesElectron Transport ChainBae SeujiNo ratings yet
- Electron Transport and Oxidative Phosphorylation: Refer To: Lehninger Principles of Biochemistry (Chapter 19)Document43 pagesElectron Transport and Oxidative Phosphorylation: Refer To: Lehninger Principles of Biochemistry (Chapter 19)Yousef KhallafNo ratings yet
- Electron TransportDocument17 pagesElectron TransportKofi Fofie-AsieduNo ratings yet
- Etc N Oxid PhoshphorylationDocument74 pagesEtc N Oxid Phoshphorylationhassanainshahi13No ratings yet
- ETC and Oxidative PhosDocument22 pagesETC and Oxidative Phos9jgdzjfpnhNo ratings yet
- 6 Electron Transport Chain NoteDocument21 pages6 Electron Transport Chain Notevgf58jsq6cNo ratings yet
- The Electron Transport ChainDocument10 pagesThe Electron Transport ChainumictNo ratings yet
- Biochem1 Fall2020 LecNov10 PostedDocument57 pagesBiochem1 Fall2020 LecNov10 PostedalexNo ratings yet
- ETC and Oxidative Phosphorylation 2021 NotesDocument9 pagesETC and Oxidative Phosphorylation 2021 NotesmunzleenkashifNo ratings yet
- 4 Bioenergetics and Oxidative Metabolism IIDocument3 pages4 Bioenergetics and Oxidative Metabolism IILinus LiuNo ratings yet
- Oxidative Phosphorylation and Electron Transport Chain II Respiratory Complexes Structure and Organization Chemiosmotic TheoryDocument20 pagesOxidative Phosphorylation and Electron Transport Chain II Respiratory Complexes Structure and Organization Chemiosmotic TheoryIffatnazNo ratings yet
- 1 Oxidative PhosphorylationDocument10 pages1 Oxidative PhosphorylationRoland ToroNo ratings yet
- Biological Oxidation: DR Nazakat Hussain Assistant ProfessorDocument32 pagesBiological Oxidation: DR Nazakat Hussain Assistant ProfessorMuhammad JamalNo ratings yet
- Biology 1: Major Features and Chemical Events of Cellular RespirationDocument5 pagesBiology 1: Major Features and Chemical Events of Cellular RespirationKaye Ann AbinalNo ratings yet
- 5-Electron Transport Chain - MDDocument31 pages5-Electron Transport Chain - MDIbrahim BarhamNo ratings yet
- How Cells Harvest EnergyDocument60 pagesHow Cells Harvest EnergyVivaMapwaNo ratings yet
- AtthapuDocument21 pagesAtthapuPrashanth PuttapagaNo ratings yet
- Online Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreDocument27 pagesOnline Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreJSlinkNYNo ratings yet
- Online Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreDocument27 pagesOnline Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreabctutorNo ratings yet
- Online Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreDocument27 pagesOnline Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreabctutorNo ratings yet
- Online Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreDocument27 pagesOnline Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreYoAmoNYC100% (1)
- Electron Transport ChainDocument11 pagesElectron Transport ChainSoofia SharifNo ratings yet
- Cellular RespirationDocument55 pagesCellular RespirationFuad sabsebNo ratings yet
- MetabolicBiochemistry Sem1 2021 Lecture6 Part2Document30 pagesMetabolicBiochemistry Sem1 2021 Lecture6 Part2kristal eliasNo ratings yet
- Plant-Physiology by TAIZ and ZEIGER PDFDocument2 pagesPlant-Physiology by TAIZ and ZEIGER PDFshxxxNo ratings yet
- Aerobic Respiration (Etc) NotesDocument11 pagesAerobic Respiration (Etc) Noteslaiyinah 09No ratings yet
- Electron Transport ChainetcDocument19 pagesElectron Transport Chainetcpk kaleenaNo ratings yet
- Respiratory Electron Transport Chain and Oxidative PhosphorylationDocument31 pagesRespiratory Electron Transport Chain and Oxidative Phosphorylationazura_zfNo ratings yet
- TCA CycleDocument22 pagesTCA CycleEMMANUEL ABEL IMAHNo ratings yet
- Electron Transport Chain andDocument25 pagesElectron Transport Chain andLovely Joy Aranda CurammengNo ratings yet
- Electron Transport ChainDocument33 pagesElectron Transport ChainAfaq AhmadNo ratings yet
- Fosforilasi OksidatifDocument82 pagesFosforilasi OksidatifSanti WilujengNo ratings yet
- Oxidative Phosphorylation and The Electron Transport Chain (ETC)Document16 pagesOxidative Phosphorylation and The Electron Transport Chain (ETC)glenn johnstonNo ratings yet
- Regulation of Oxidative PhosphorylationDocument14 pagesRegulation of Oxidative Phosphorylationmaaz629No ratings yet
- Electron Transport Chain: Jump To Navigation Jump To SearchDocument16 pagesElectron Transport Chain: Jump To Navigation Jump To SearchJennie KimNo ratings yet
- BiochemDocument10 pagesBiochemHoàng LâmNo ratings yet
- 2 Lec. Biochemistry (4th)Document25 pages2 Lec. Biochemistry (4th)Doctor SonuNo ratings yet
- Electron Transport ChainDocument7 pagesElectron Transport ChainMaria HarisNo ratings yet
- Ch. 9 Biological OxidationDocument71 pagesCh. 9 Biological OxidationKrishna KanthNo ratings yet
- In Relation To Lsm2101 Lec 2Document11 pagesIn Relation To Lsm2101 Lec 2jojolim18No ratings yet
- Chapter 20 ETC and Oxidative PhosphDocument29 pagesChapter 20 ETC and Oxidative PhosphSpencer ThomasNo ratings yet
- Electron Transport ChainDocument11 pagesElectron Transport ChainsakshibhandariNo ratings yet
- New Frontiers in Sciences, Engineering and the Arts: Vol. Ii the Chemistry of Initiation of Non-Ringed Monomers/CompoundsFrom EverandNew Frontiers in Sciences, Engineering and the Arts: Vol. Ii the Chemistry of Initiation of Non-Ringed Monomers/CompoundsNo ratings yet
- Title: Haem Metabolism: Porphyrias: Name: DR Yemisi Latunde-DadaDocument37 pagesTitle: Haem Metabolism: Porphyrias: Name: DR Yemisi Latunde-DadaW BNo ratings yet
- Carbohydrate Metab - Lectures 3,4 Slides, 2020-21Document15 pagesCarbohydrate Metab - Lectures 3,4 Slides, 2020-21W BNo ratings yet
- 5BBB0223: Carbohydrates and Metabolism: Peter EllisDocument14 pages5BBB0223: Carbohydrates and Metabolism: Peter EllisW BNo ratings yet
- 5BBB0223 Metabolic Integration PSW1 ExplanationDocument25 pages5BBB0223 Metabolic Integration PSW1 ExplanationW BNo ratings yet
- Acids Answer ChemistryDocument1 pageAcids Answer ChemistryW BNo ratings yet
- 5BBB0223 TBL2 - Mitochondria: Answer SheetDocument2 pages5BBB0223 TBL2 - Mitochondria: Answer SheetW BNo ratings yet
- 5BBB0223 TBL2 - MitochondriaDocument2 pages5BBB0223 TBL2 - MitochondriaW BNo ratings yet
- Chemistry - Qualitative AnalysisDocument2 pagesChemistry - Qualitative AnalysisAditya SenthilNo ratings yet
- BioplasticDocument33 pagesBioplasticCatherine RajanNo ratings yet
- 5243 Heterogeneous Catalysis1Document7 pages5243 Heterogeneous Catalysis1Mohit PatelNo ratings yet
- Vishnupriya Cs MoocDocument16 pagesVishnupriya Cs MoocJyothy ShajiNo ratings yet
- Unit 3 Lipids Lesson 3: The Cell MembraneDocument12 pagesUnit 3 Lipids Lesson 3: The Cell MembraneValenzuela Allene GraceNo ratings yet
- Experiment 1A and 1BDocument8 pagesExperiment 1A and 1BManda BaboolalNo ratings yet
- Jam Processing - Effect of Pectin Replacement by Locust Bean Gum On Its CharacteristicsDocument12 pagesJam Processing - Effect of Pectin Replacement by Locust Bean Gum On Its CharacteristicsThảo PhạmNo ratings yet
- Exam Final SugralionaDocument1 pageExam Final SugralionaDaniyar TemirovNo ratings yet
- Production of Yarn From PlasticDocument11 pagesProduction of Yarn From PlasticS.R.ANEESHNo ratings yet
- Biology Model Exam For G-12 2016 Addis KetemaDocument12 pagesBiology Model Exam For G-12 2016 Addis Ketemamuller.eshetu12No ratings yet
- Waukesha Digester GasDocument13 pagesWaukesha Digester GasA2 BhaskarNo ratings yet
- 9700 BIOLOGY: MARK SCHEME For The October/November 2015 SeriesDocument9 pages9700 BIOLOGY: MARK SCHEME For The October/November 2015 SeriesNerminNo ratings yet
- Literature Review MitragynineDocument8 pagesLiterature Review MitragynineMuallif HasanNo ratings yet
- NB DhavalDocument140 pagesNB DhavalDEVESH SINGH100% (2)
- Time: 1 Hrs Max. Marks: 98 Single Correct: 3 2 4 HG ZNDocument6 pagesTime: 1 Hrs Max. Marks: 98 Single Correct: 3 2 4 HG ZNlakshmi.vedanarayanan7785No ratings yet
- Sri Chaitanya Chemistry MULTIPLE CORRECT Type Question BANKkkkkkkkkkkkkkkkkkkkkkkkkkkkDocument36 pagesSri Chaitanya Chemistry MULTIPLE CORRECT Type Question BANKkkkkkkkkkkkkkkkkkkkkkkkkkkkSai Gokul0% (1)
- Ra 6969Document61 pagesRa 6969Charles Rommel TadoNo ratings yet
- 301 Chemistry Model Question Paper New CourseDocument9 pages301 Chemistry Model Question Paper New CourseZhu ZhuNo ratings yet
- Lesson Plan Protein SynthesisDocument2 pagesLesson Plan Protein SynthesisQueencess Ara Torres100% (1)
- (Encoded Version) Q1 - WS - Lesson4 - Nomenclature of Inorganic Compounds V2Document6 pages(Encoded Version) Q1 - WS - Lesson4 - Nomenclature of Inorganic Compounds V2Leslie BenetezNo ratings yet
- Pharmacognostic, Phytochemical and in Vivo Hepatoprotective Activity On Pongamia Pinnata Linn BarkDocument10 pagesPharmacognostic, Phytochemical and in Vivo Hepatoprotective Activity On Pongamia Pinnata Linn BarkDr. Ramadevi DevarakondaNo ratings yet
- Alkenes: Structure HDocument8 pagesAlkenes: Structure HOkeileo OsbourneNo ratings yet
- Product Data Sheet: Eastman™ Cellulose Acetate Butyrate (CAB-551-0.2) Application/UsesDocument2 pagesProduct Data Sheet: Eastman™ Cellulose Acetate Butyrate (CAB-551-0.2) Application/UsesBERKAT RAJA SEMESTANo ratings yet
- Peroxyacetyl Nitrate (Pan) : Historical PerspectiveDocument8 pagesPeroxyacetyl Nitrate (Pan) : Historical PerspectivehjbhhbjNo ratings yet
- A Review On Edible Straw Mushrooms: A Source of High Nutritional Supplement, Biologically Active Diverse Structural PolysaccharidesDocument10 pagesA Review On Edible Straw Mushrooms: A Source of High Nutritional Supplement, Biologically Active Diverse Structural PolysaccharidesViedya YadaNo ratings yet
- ICNF2023 Program DAY1Document4 pagesICNF2023 Program DAY1Vijay BahetiNo ratings yet
- Exhaust SystemDocument16 pagesExhaust SystemArvind KatyayanNo ratings yet
- PHD Thesis Yale UniversityDocument4 pagesPHD Thesis Yale Universitycarlabenningtonnaperville100% (2)
- Batch 14Document22 pagesBatch 14Sreenivas NaruNo ratings yet