
Cereb. Cortex-2009-LoTurco-i120-5
Cereb. Cortex-2009-LoTurco-i120-5
You might also like
- Lab Manual in Genetics 2019Document125 pagesLab Manual in Genetics 2019max delvalle100% (5)
- Diphtheria Schematic Diagram (Pathophysiology0Document3 pagesDiphtheria Schematic Diagram (Pathophysiology0Kathlene Boleche100% (1)
- Direct Visualization of Nucleogenesis by Precerebellar Neurons: Involvement of Ventricle-Directed, Radial Fibre-Associated MigrationDocument11 pagesDirect Visualization of Nucleogenesis by Precerebellar Neurons: Involvement of Ventricle-Directed, Radial Fibre-Associated Migrationhareb_dNo ratings yet
- Des Car GableDocument10 pagesDes Car GableDago CastilloNo ratings yet
- Magnetic Resonance Imaging of Iron Oxide Labelled Stem Cells: Applications To Tissue Engineering Based Regeneration of The Intervertebral DiscDocument9 pagesMagnetic Resonance Imaging of Iron Oxide Labelled Stem Cells: Applications To Tissue Engineering Based Regeneration of The Intervertebral DiscArdhina BeliberzNo ratings yet
- The Drosophila JNK Pathway Controls The Morphogenesis of The Egg Dorsal Appendages and MicropyleDocument13 pagesThe Drosophila JNK Pathway Controls The Morphogenesis of The Egg Dorsal Appendages and MicropyleFernandoNo ratings yet
- BIO99 Cellular Reprogramming - Pluripotent Stem Cells (Yamanaka)Document11 pagesBIO99 Cellular Reprogramming - Pluripotent Stem Cells (Yamanaka)Paolo GochingcoNo ratings yet
- Cell Surface Fluctuations Regulate Early Embryonic Lineage SortingDocument38 pagesCell Surface Fluctuations Regulate Early Embryonic Lineage SortingFenrir1017No ratings yet
- Efficient Targeting of Gene Expression in Chick Embryos by MicroelectroporationDocument10 pagesEfficient Targeting of Gene Expression in Chick Embryos by MicroelectroporationRaynNo ratings yet
- Spatiotemporal Molecular Approach of in Utero Electroporation To Functionally Decipher Endophenotypes in Neurodevelopmental DisordersDocument7 pagesSpatiotemporal Molecular Approach of in Utero Electroporation To Functionally Decipher Endophenotypes in Neurodevelopmental DisordersAnnetrude de MooijNo ratings yet
- 2009-High Resolution Map of Caenorhabditis Elegans Gap Junction ProteinsDocument15 pages2009-High Resolution Map of Caenorhabditis Elegans Gap Junction ProteinsyicinenNo ratings yet
- 2009-High Resolution Map of Caenorhabditis Elegans Gap Junction ProteinsDocument15 pages2009-High Resolution Map of Caenorhabditis Elegans Gap Junction ProteinsyicinenNo ratings yet
- Encyclopediabookchapter PDFDocument13 pagesEncyclopediabookchapter PDFJamesNo ratings yet
- Inexhaustible Hair-Cell Regeneration in Young and Aged ZebrafishDocument7 pagesInexhaustible Hair-Cell Regeneration in Young and Aged ZebrafishSaifuddin HaswareNo ratings yet
- Ni Hms 393652Document6 pagesNi Hms 39365222gaefNo ratings yet
- A Transcriptional LogicDocument4 pagesA Transcriptional LogicMaximilian PfauNo ratings yet
- Jacobs, Ku, Que - 2012 - Genetic and Cellular Mechanisms Regulating Anterior Foregut and Esophageal DevelopmentDocument11 pagesJacobs, Ku, Que - 2012 - Genetic and Cellular Mechanisms Regulating Anterior Foregut and Esophageal DevelopmentCarly LeungNo ratings yet
- TRKB Downregulation Is Required For Dendrite Retraction in Developing Neurons of Chicken Nucleus MagnocellularisDocument10 pagesTRKB Downregulation Is Required For Dendrite Retraction in Developing Neurons of Chicken Nucleus MagnocellularisFrontiersNo ratings yet
- A Drosophila Gain-of-Function Screen For Candidate Genes Involved in Steroid-Dependent Neuroendocrine Cell RemodelingDocument19 pagesA Drosophila Gain-of-Function Screen For Candidate Genes Involved in Steroid-Dependent Neuroendocrine Cell RemodelingLautaro BelfioriNo ratings yet
- Tracking in Vitro and in Vivo Sirna Electrotransfer in Tumor CellsDocument8 pagesTracking in Vitro and in Vivo Sirna Electrotransfer in Tumor Cellscindy tatiana rodriguez marinNo ratings yet
- Developmento of Vestibular SystemDocument11 pagesDevelopmento of Vestibular Systempaula.rhytmosNo ratings yet
- Clinical Potentials of Human Pluripotent Stem CellsDocument10 pagesClinical Potentials of Human Pluripotent Stem CellsMauricio ChamorroNo ratings yet
- Boquila TrifoliataDocument3 pagesBoquila Trifoliataiperico68No ratings yet
- Vision in Plants Via Plant-Specific Ocelli?: ForumDocument4 pagesVision in Plants Via Plant-Specific Ocelli?: ForumEmmanuel GrimaudNo ratings yet
- TMP 6 DAEDocument16 pagesTMP 6 DAEFrontiersNo ratings yet
- Paper Canty 2009 - PV HolesDocument11 pagesPaper Canty 2009 - PV Holesmaariamunoz9No ratings yet
- Integration of Word Meaning and World Knowledge in Language ComprehensionDocument5 pagesIntegration of Word Meaning and World Knowledge in Language ComprehensionKarina AnnesNo ratings yet
- ICMto ESCDocument11 pagesICMto ESCzotzedNo ratings yet
- Nuclear Pore ComplexesDocument9 pagesNuclear Pore ComplexesMNo ratings yet
- (Review) (Biomolecular Concepts) Kazuyuki Ohbo Et Shin-Ichi Tomizawa-2015Document9 pages(Review) (Biomolecular Concepts) Kazuyuki Ohbo Et Shin-Ichi Tomizawa-2015Priscilla FreschuNo ratings yet
- (Vincent Et Al, 1997) Ntegration of The Head and Trunk Segmentation Systems Controls Cephalic Furrow Formation in DrosophilaDocument8 pages(Vincent Et Al, 1997) Ntegration of The Head and Trunk Segmentation Systems Controls Cephalic Furrow Formation in DrosophilaエリタノアンソネーNo ratings yet
- Special Review Series - Gene Manipulation and Integrative PhysiologyDocument13 pagesSpecial Review Series - Gene Manipulation and Integrative PhysiologyAnunay JoshiNo ratings yet
- Review Induction of Pluripotency by de Fined Factors: Amanaka M.J.ADocument14 pagesReview Induction of Pluripotency by de Fined Factors: Amanaka M.J.AnembutalNo ratings yet
- Tribolium Segementation Clock 2012 El Sherif Et AlDocument6 pagesTribolium Segementation Clock 2012 El Sherif Et AlCarmen CoelhoNo ratings yet
- FT 1329Document11 pagesFT 1329aaki2627No ratings yet
- Micro e Nanoestrutura CelularDocument4 pagesMicro e Nanoestrutura CelularIsis CôrtesNo ratings yet
- The Use of Zebrafish To ReviewDocument13 pagesThe Use of Zebrafish To ReviewKarthik ShettyNo ratings yet
- IKNMDocument31 pagesIKNMray goldsteinNo ratings yet
- Self-Organized Nuclear Positioning Synchronizes The Cell Cycle in Drosophila EmbryosDocument35 pagesSelf-Organized Nuclear Positioning Synchronizes The Cell Cycle in Drosophila EmbryosエリタノアンソネーNo ratings yet
- Electropermeabilization, A Physical Method For The Delivery of Therapeutic Molecules Into CellsDocument6 pagesElectropermeabilization, A Physical Method For The Delivery of Therapeutic Molecules Into CellsEspacio UniversitarioNo ratings yet
- Advances in Reprogramming Somatic Cells To InducedDocument21 pagesAdvances in Reprogramming Somatic Cells To InducedNatalia MoysésNo ratings yet
- Huang2012 PDFDocument13 pagesHuang2012 PDFtsth4ckNo ratings yet
- Flpstop, A Tool For Conditional Gene Control in DrosophilaDocument33 pagesFlpstop, A Tool For Conditional Gene Control in DrosophilahaihuaiheNo ratings yet
- Royer 2020Document12 pagesRoyer 2020nugrahoneyNo ratings yet
- Astrocytes Biology and PathologyDocument29 pagesAstrocytes Biology and PathologyCristina SanchezNo ratings yet
- Self-Organizing Properties of Mouse Pluripotent Cells Initiate Morphogenesis Upon ImplantationDocument18 pagesSelf-Organizing Properties of Mouse Pluripotent Cells Initiate Morphogenesis Upon Implantationiakyol19No ratings yet
- Thalamic Neuron TheoryDocument18 pagesThalamic Neuron TheorybazediNo ratings yet
- FullDocument8 pagesFullrocambolescas perthNo ratings yet
- Genes Dev. 2015 Quesnel Vallières 746 59Document15 pagesGenes Dev. 2015 Quesnel Vallières 746 59Misael G JordanNo ratings yet
- The Mouse Epididymal Transcriptome: Transcriptional Profiling of Segmental Gene Expression in The EpididymisDocument10 pagesThe Mouse Epididymal Transcriptome: Transcriptional Profiling of Segmental Gene Expression in The EpididymisAhmad SolihinNo ratings yet
- Genome Organization and Function: A View From Yeast and ArabidopsisDocument13 pagesGenome Organization and Function: A View From Yeast and ArabidopsisJulienne InostrozaNo ratings yet
- X ENEURO.0337-20.2021.fullDocument24 pagesX ENEURO.0337-20.2021.fullloherjulian6No ratings yet
- Integrin-Linked Kinase Deletion From Mouse Cortex Results in Cortical Lamination Defects Resembling Cobblestone LissencephalyDocument10 pagesIntegrin-Linked Kinase Deletion From Mouse Cortex Results in Cortical Lamination Defects Resembling Cobblestone LissencephalyCris AdFNo ratings yet
- Learn. Mem.-2009-Jessberger-147-54llDocument9 pagesLearn. Mem.-2009-Jessberger-147-54llJean Pierre Chastre LuzaNo ratings yet
- JIPB - 2014 - Hawes - The Endoplasmic Reticulum A Dynamic and Well Connected OrganelleDocument13 pagesJIPB - 2014 - Hawes - The Endoplasmic Reticulum A Dynamic and Well Connected Organellekaviyarajkumar6No ratings yet
- 1644 FullDocument5 pages1644 FullhermannNo ratings yet
- Order 336296Document11 pagesOrder 336296Isaac Wachira MuneneNo ratings yet
- The Common NF-κB Essential Modulator (NEMO) Gene Rearrangement in Korean Patients with Incontinentia PigmentiDocument6 pagesThe Common NF-κB Essential Modulator (NEMO) Gene Rearrangement in Korean Patients with Incontinentia PigmentiAriefBayuArdiantoNo ratings yet
- J Bbagrm 2009 02 002Document7 pagesJ Bbagrm 2009 02 002Raymundo Lopez NNo ratings yet
- Edidin 202003Document6 pagesEdidin 202003david torresNo ratings yet
- INsights Into Protein SynthesisDocument9 pagesINsights Into Protein SynthesisRam KNo ratings yet
- THE LOBSTER:: a Model for Teaching Neurophysiological ConceptsFrom EverandTHE LOBSTER:: a Model for Teaching Neurophysiological ConceptsNo ratings yet
- 337d PDFDocument8 pages337d PDFEka MeylianaNo ratings yet
- Acute Frontal Sinusitis - Sics EditoreDocument49 pagesAcute Frontal Sinusitis - Sics Editoreslowgrind9719No ratings yet
- Table 2A Enterobacterales M02 and M07Document2 pagesTable 2A Enterobacterales M02 and M07helenmariana.ramosNo ratings yet
- Chapter Protein Folding and MisfoldingDocument17 pagesChapter Protein Folding and MisfoldingMazhar FarNo ratings yet
- DNA ReplicationDocument2 pagesDNA ReplicationFrancoise AngeliqueNo ratings yet
- Inflammation, Infection, Immunity, and Wound Care - NCLEX - SECEDocument29 pagesInflammation, Infection, Immunity, and Wound Care - NCLEX - SECENadesNo ratings yet
- Biological ClasifficationDocument6 pagesBiological Clasifficationchannel of dhyanaNo ratings yet
- Fallone 2019Document10 pagesFallone 2019Noel Saúl Argüello SánchezNo ratings yet
- FYBSc Sem-I Paper-II PDFDocument26 pagesFYBSc Sem-I Paper-II PDFbjkopparNo ratings yet
- Final Istb 2016Document192 pagesFinal Istb 2016yudhaputraNo ratings yet
- 5 HypersensitivityDocument13 pages5 HypersensitivityPurityNo ratings yet
- Gram StainDocument3 pagesGram StainAbduladheemNo ratings yet
- Mono - Probiotics - English MONOGRAFIA HEALTH CANADA - 0Document25 pagesMono - Probiotics - English MONOGRAFIA HEALTH CANADA - 0Farhan aliNo ratings yet
- Immunity and Vaccinations 13/11/2019: Complete Your Now Try This!Document25 pagesImmunity and Vaccinations 13/11/2019: Complete Your Now Try This!sai yadaNo ratings yet
- WK13 CDN - Meningitis, Poliomyelitis, Leprosy, Tetanus, Red TideDocument43 pagesWK13 CDN - Meningitis, Poliomyelitis, Leprosy, Tetanus, Red TidePotato PceeNo ratings yet
- 1 s2.0 S1934590920302009 MainDocument15 pages1 s2.0 S1934590920302009 MainMOVEMENT SCHOOLNo ratings yet
- 5e Plan-Central DogmaDocument2 pages5e Plan-Central Dogmaapi-307962259No ratings yet
- S807 Mini Review v4Document37 pagesS807 Mini Review v4Daen de LeonNo ratings yet
- Lesson Plan 7 FungiDocument4 pagesLesson Plan 7 FungiLarraine Bernice Gannaban Cabildo100% (9)
- Feline Panleukopenia Balram YadavDocument16 pagesFeline Panleukopenia Balram Yadavsanchi ranaNo ratings yet
- Types of PneumoniaDocument6 pagesTypes of PneumoniasakuraleeshaoranNo ratings yet
- Chapter 5-Obstetrics Conditions Affects The NeonateDocument129 pagesChapter 5-Obstetrics Conditions Affects The Neonatesinte beyuNo ratings yet
- A Retrospective Study of Poultry Diseases Diagnosed in Maiduguri Northeast Nigeria pfw.1000113 PDFDocument4 pagesA Retrospective Study of Poultry Diseases Diagnosed in Maiduguri Northeast Nigeria pfw.1000113 PDFWahyu HidayatNo ratings yet
- Bio-Energetic Frequency ChartsDocument37 pagesBio-Energetic Frequency ChartsHayden JamesNo ratings yet
- Sciencedirect Food Fermentations For Improved Digestibility of Plant Foods - An Essential Ex Situ Digestion Step in Agricultural Societies?Document8 pagesSciencedirect Food Fermentations For Improved Digestibility of Plant Foods - An Essential Ex Situ Digestion Step in Agricultural Societies?arNo ratings yet
- Antimicrobial ResistanceDocument4 pagesAntimicrobial ResistanceDip Ayan MNo ratings yet
- Pathology+101 Complete)Document147 pagesPathology+101 Complete)Goh Kah Yong100% (2)
- Study Guide For Module No. - : Cytological Bases of HeredityDocument12 pagesStudy Guide For Module No. - : Cytological Bases of HeredityREGINA MAE JUNIONo ratings yet

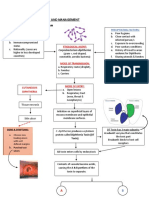























































































Download as pdf or txt
You might also like
- Lab Manual in Genetics 2019Document125 pagesLab Manual in Genetics 2019max delvalle100% (5)
- Diphtheria Schematic Diagram (Pathophysiology0Document3 pagesDiphtheria Schematic Diagram (Pathophysiology0Kathlene Boleche100% (1)
- Direct Visualization of Nucleogenesis by Precerebellar Neurons: Involvement of Ventricle-Directed, Radial Fibre-Associated MigrationDocument11 pagesDirect Visualization of Nucleogenesis by Precerebellar Neurons: Involvement of Ventricle-Directed, Radial Fibre-Associated Migrationhareb_dNo ratings yet
- Des Car GableDocument10 pagesDes Car GableDago CastilloNo ratings yet
- Magnetic Resonance Imaging of Iron Oxide Labelled Stem Cells: Applications To Tissue Engineering Based Regeneration of The Intervertebral DiscDocument9 pagesMagnetic Resonance Imaging of Iron Oxide Labelled Stem Cells: Applications To Tissue Engineering Based Regeneration of The Intervertebral DiscArdhina BeliberzNo ratings yet
- The Drosophila JNK Pathway Controls The Morphogenesis of The Egg Dorsal Appendages and MicropyleDocument13 pagesThe Drosophila JNK Pathway Controls The Morphogenesis of The Egg Dorsal Appendages and MicropyleFernandoNo ratings yet
- BIO99 Cellular Reprogramming - Pluripotent Stem Cells (Yamanaka)Document11 pagesBIO99 Cellular Reprogramming - Pluripotent Stem Cells (Yamanaka)Paolo GochingcoNo ratings yet
- Cell Surface Fluctuations Regulate Early Embryonic Lineage SortingDocument38 pagesCell Surface Fluctuations Regulate Early Embryonic Lineage SortingFenrir1017No ratings yet
- Efficient Targeting of Gene Expression in Chick Embryos by MicroelectroporationDocument10 pagesEfficient Targeting of Gene Expression in Chick Embryos by MicroelectroporationRaynNo ratings yet
- Spatiotemporal Molecular Approach of in Utero Electroporation To Functionally Decipher Endophenotypes in Neurodevelopmental DisordersDocument7 pagesSpatiotemporal Molecular Approach of in Utero Electroporation To Functionally Decipher Endophenotypes in Neurodevelopmental DisordersAnnetrude de MooijNo ratings yet
- 2009-High Resolution Map of Caenorhabditis Elegans Gap Junction ProteinsDocument15 pages2009-High Resolution Map of Caenorhabditis Elegans Gap Junction ProteinsyicinenNo ratings yet
- 2009-High Resolution Map of Caenorhabditis Elegans Gap Junction ProteinsDocument15 pages2009-High Resolution Map of Caenorhabditis Elegans Gap Junction ProteinsyicinenNo ratings yet
- Encyclopediabookchapter PDFDocument13 pagesEncyclopediabookchapter PDFJamesNo ratings yet
- Inexhaustible Hair-Cell Regeneration in Young and Aged ZebrafishDocument7 pagesInexhaustible Hair-Cell Regeneration in Young and Aged ZebrafishSaifuddin HaswareNo ratings yet
- Ni Hms 393652Document6 pagesNi Hms 39365222gaefNo ratings yet
- A Transcriptional LogicDocument4 pagesA Transcriptional LogicMaximilian PfauNo ratings yet
- Jacobs, Ku, Que - 2012 - Genetic and Cellular Mechanisms Regulating Anterior Foregut and Esophageal DevelopmentDocument11 pagesJacobs, Ku, Que - 2012 - Genetic and Cellular Mechanisms Regulating Anterior Foregut and Esophageal DevelopmentCarly LeungNo ratings yet
- TRKB Downregulation Is Required For Dendrite Retraction in Developing Neurons of Chicken Nucleus MagnocellularisDocument10 pagesTRKB Downregulation Is Required For Dendrite Retraction in Developing Neurons of Chicken Nucleus MagnocellularisFrontiersNo ratings yet
- A Drosophila Gain-of-Function Screen For Candidate Genes Involved in Steroid-Dependent Neuroendocrine Cell RemodelingDocument19 pagesA Drosophila Gain-of-Function Screen For Candidate Genes Involved in Steroid-Dependent Neuroendocrine Cell RemodelingLautaro BelfioriNo ratings yet
- Tracking in Vitro and in Vivo Sirna Electrotransfer in Tumor CellsDocument8 pagesTracking in Vitro and in Vivo Sirna Electrotransfer in Tumor Cellscindy tatiana rodriguez marinNo ratings yet
- Developmento of Vestibular SystemDocument11 pagesDevelopmento of Vestibular Systempaula.rhytmosNo ratings yet
- Clinical Potentials of Human Pluripotent Stem CellsDocument10 pagesClinical Potentials of Human Pluripotent Stem CellsMauricio ChamorroNo ratings yet
- Boquila TrifoliataDocument3 pagesBoquila Trifoliataiperico68No ratings yet
- Vision in Plants Via Plant-Specific Ocelli?: ForumDocument4 pagesVision in Plants Via Plant-Specific Ocelli?: ForumEmmanuel GrimaudNo ratings yet
- TMP 6 DAEDocument16 pagesTMP 6 DAEFrontiersNo ratings yet
- Paper Canty 2009 - PV HolesDocument11 pagesPaper Canty 2009 - PV Holesmaariamunoz9No ratings yet
- Integration of Word Meaning and World Knowledge in Language ComprehensionDocument5 pagesIntegration of Word Meaning and World Knowledge in Language ComprehensionKarina AnnesNo ratings yet
- ICMto ESCDocument11 pagesICMto ESCzotzedNo ratings yet
- Nuclear Pore ComplexesDocument9 pagesNuclear Pore ComplexesMNo ratings yet
- (Review) (Biomolecular Concepts) Kazuyuki Ohbo Et Shin-Ichi Tomizawa-2015Document9 pages(Review) (Biomolecular Concepts) Kazuyuki Ohbo Et Shin-Ichi Tomizawa-2015Priscilla FreschuNo ratings yet
- (Vincent Et Al, 1997) Ntegration of The Head and Trunk Segmentation Systems Controls Cephalic Furrow Formation in DrosophilaDocument8 pages(Vincent Et Al, 1997) Ntegration of The Head and Trunk Segmentation Systems Controls Cephalic Furrow Formation in DrosophilaエリタノアンソネーNo ratings yet
- Special Review Series - Gene Manipulation and Integrative PhysiologyDocument13 pagesSpecial Review Series - Gene Manipulation and Integrative PhysiologyAnunay JoshiNo ratings yet
- Review Induction of Pluripotency by de Fined Factors: Amanaka M.J.ADocument14 pagesReview Induction of Pluripotency by de Fined Factors: Amanaka M.J.AnembutalNo ratings yet
- Tribolium Segementation Clock 2012 El Sherif Et AlDocument6 pagesTribolium Segementation Clock 2012 El Sherif Et AlCarmen CoelhoNo ratings yet
- FT 1329Document11 pagesFT 1329aaki2627No ratings yet
- Micro e Nanoestrutura CelularDocument4 pagesMicro e Nanoestrutura CelularIsis CôrtesNo ratings yet
- The Use of Zebrafish To ReviewDocument13 pagesThe Use of Zebrafish To ReviewKarthik ShettyNo ratings yet
- IKNMDocument31 pagesIKNMray goldsteinNo ratings yet
- Self-Organized Nuclear Positioning Synchronizes The Cell Cycle in Drosophila EmbryosDocument35 pagesSelf-Organized Nuclear Positioning Synchronizes The Cell Cycle in Drosophila EmbryosエリタノアンソネーNo ratings yet
- Electropermeabilization, A Physical Method For The Delivery of Therapeutic Molecules Into CellsDocument6 pagesElectropermeabilization, A Physical Method For The Delivery of Therapeutic Molecules Into CellsEspacio UniversitarioNo ratings yet
- Advances in Reprogramming Somatic Cells To InducedDocument21 pagesAdvances in Reprogramming Somatic Cells To InducedNatalia MoysésNo ratings yet
- Huang2012 PDFDocument13 pagesHuang2012 PDFtsth4ckNo ratings yet
- Flpstop, A Tool For Conditional Gene Control in DrosophilaDocument33 pagesFlpstop, A Tool For Conditional Gene Control in DrosophilahaihuaiheNo ratings yet
- Royer 2020Document12 pagesRoyer 2020nugrahoneyNo ratings yet
- Astrocytes Biology and PathologyDocument29 pagesAstrocytes Biology and PathologyCristina SanchezNo ratings yet
- Self-Organizing Properties of Mouse Pluripotent Cells Initiate Morphogenesis Upon ImplantationDocument18 pagesSelf-Organizing Properties of Mouse Pluripotent Cells Initiate Morphogenesis Upon Implantationiakyol19No ratings yet
- Thalamic Neuron TheoryDocument18 pagesThalamic Neuron TheorybazediNo ratings yet
- FullDocument8 pagesFullrocambolescas perthNo ratings yet
- Genes Dev. 2015 Quesnel Vallières 746 59Document15 pagesGenes Dev. 2015 Quesnel Vallières 746 59Misael G JordanNo ratings yet
- The Mouse Epididymal Transcriptome: Transcriptional Profiling of Segmental Gene Expression in The EpididymisDocument10 pagesThe Mouse Epididymal Transcriptome: Transcriptional Profiling of Segmental Gene Expression in The EpididymisAhmad SolihinNo ratings yet
- Genome Organization and Function: A View From Yeast and ArabidopsisDocument13 pagesGenome Organization and Function: A View From Yeast and ArabidopsisJulienne InostrozaNo ratings yet
- X ENEURO.0337-20.2021.fullDocument24 pagesX ENEURO.0337-20.2021.fullloherjulian6No ratings yet
- Integrin-Linked Kinase Deletion From Mouse Cortex Results in Cortical Lamination Defects Resembling Cobblestone LissencephalyDocument10 pagesIntegrin-Linked Kinase Deletion From Mouse Cortex Results in Cortical Lamination Defects Resembling Cobblestone LissencephalyCris AdFNo ratings yet
- Learn. Mem.-2009-Jessberger-147-54llDocument9 pagesLearn. Mem.-2009-Jessberger-147-54llJean Pierre Chastre LuzaNo ratings yet
- JIPB - 2014 - Hawes - The Endoplasmic Reticulum A Dynamic and Well Connected OrganelleDocument13 pagesJIPB - 2014 - Hawes - The Endoplasmic Reticulum A Dynamic and Well Connected Organellekaviyarajkumar6No ratings yet
- 1644 FullDocument5 pages1644 FullhermannNo ratings yet
- Order 336296Document11 pagesOrder 336296Isaac Wachira MuneneNo ratings yet
- The Common NF-κB Essential Modulator (NEMO) Gene Rearrangement in Korean Patients with Incontinentia PigmentiDocument6 pagesThe Common NF-κB Essential Modulator (NEMO) Gene Rearrangement in Korean Patients with Incontinentia PigmentiAriefBayuArdiantoNo ratings yet
- J Bbagrm 2009 02 002Document7 pagesJ Bbagrm 2009 02 002Raymundo Lopez NNo ratings yet
- Edidin 202003Document6 pagesEdidin 202003david torresNo ratings yet
- INsights Into Protein SynthesisDocument9 pagesINsights Into Protein SynthesisRam KNo ratings yet
- THE LOBSTER:: a Model for Teaching Neurophysiological ConceptsFrom EverandTHE LOBSTER:: a Model for Teaching Neurophysiological ConceptsNo ratings yet
- 337d PDFDocument8 pages337d PDFEka MeylianaNo ratings yet
- Acute Frontal Sinusitis - Sics EditoreDocument49 pagesAcute Frontal Sinusitis - Sics Editoreslowgrind9719No ratings yet
- Table 2A Enterobacterales M02 and M07Document2 pagesTable 2A Enterobacterales M02 and M07helenmariana.ramosNo ratings yet
- Chapter Protein Folding and MisfoldingDocument17 pagesChapter Protein Folding and MisfoldingMazhar FarNo ratings yet
- DNA ReplicationDocument2 pagesDNA ReplicationFrancoise AngeliqueNo ratings yet
- Inflammation, Infection, Immunity, and Wound Care - NCLEX - SECEDocument29 pagesInflammation, Infection, Immunity, and Wound Care - NCLEX - SECENadesNo ratings yet
- Biological ClasifficationDocument6 pagesBiological Clasifficationchannel of dhyanaNo ratings yet
- Fallone 2019Document10 pagesFallone 2019Noel Saúl Argüello SánchezNo ratings yet
- FYBSc Sem-I Paper-II PDFDocument26 pagesFYBSc Sem-I Paper-II PDFbjkopparNo ratings yet
- Final Istb 2016Document192 pagesFinal Istb 2016yudhaputraNo ratings yet
- 5 HypersensitivityDocument13 pages5 HypersensitivityPurityNo ratings yet
- Gram StainDocument3 pagesGram StainAbduladheemNo ratings yet
- Mono - Probiotics - English MONOGRAFIA HEALTH CANADA - 0Document25 pagesMono - Probiotics - English MONOGRAFIA HEALTH CANADA - 0Farhan aliNo ratings yet
- Immunity and Vaccinations 13/11/2019: Complete Your Now Try This!Document25 pagesImmunity and Vaccinations 13/11/2019: Complete Your Now Try This!sai yadaNo ratings yet
- WK13 CDN - Meningitis, Poliomyelitis, Leprosy, Tetanus, Red TideDocument43 pagesWK13 CDN - Meningitis, Poliomyelitis, Leprosy, Tetanus, Red TidePotato PceeNo ratings yet
- 1 s2.0 S1934590920302009 MainDocument15 pages1 s2.0 S1934590920302009 MainMOVEMENT SCHOOLNo ratings yet
- 5e Plan-Central DogmaDocument2 pages5e Plan-Central Dogmaapi-307962259No ratings yet
- S807 Mini Review v4Document37 pagesS807 Mini Review v4Daen de LeonNo ratings yet
- Lesson Plan 7 FungiDocument4 pagesLesson Plan 7 FungiLarraine Bernice Gannaban Cabildo100% (9)
- Feline Panleukopenia Balram YadavDocument16 pagesFeline Panleukopenia Balram Yadavsanchi ranaNo ratings yet
- Types of PneumoniaDocument6 pagesTypes of PneumoniasakuraleeshaoranNo ratings yet
- Chapter 5-Obstetrics Conditions Affects The NeonateDocument129 pagesChapter 5-Obstetrics Conditions Affects The Neonatesinte beyuNo ratings yet
- A Retrospective Study of Poultry Diseases Diagnosed in Maiduguri Northeast Nigeria pfw.1000113 PDFDocument4 pagesA Retrospective Study of Poultry Diseases Diagnosed in Maiduguri Northeast Nigeria pfw.1000113 PDFWahyu HidayatNo ratings yet
- Bio-Energetic Frequency ChartsDocument37 pagesBio-Energetic Frequency ChartsHayden JamesNo ratings yet
- Sciencedirect Food Fermentations For Improved Digestibility of Plant Foods - An Essential Ex Situ Digestion Step in Agricultural Societies?Document8 pagesSciencedirect Food Fermentations For Improved Digestibility of Plant Foods - An Essential Ex Situ Digestion Step in Agricultural Societies?arNo ratings yet
- Antimicrobial ResistanceDocument4 pagesAntimicrobial ResistanceDip Ayan MNo ratings yet
- Pathology+101 Complete)Document147 pagesPathology+101 Complete)Goh Kah Yong100% (2)
- Study Guide For Module No. - : Cytological Bases of HeredityDocument12 pagesStudy Guide For Module No. - : Cytological Bases of HeredityREGINA MAE JUNIONo ratings yet